
基于水质浮标在线监测的米氏凯伦藻赤潮过程及环境因子变化特征分析
作者简介:赵聪蛟(1981—), 男, 河北省柏乡县人, 工程师, 硕士, 主要从事海洋浮标、湿地和海洋生态研究与评价。E-mail: congcong990@163.com
*感谢两位匿名审稿人提出宝贵的意见。
Editor: 殷波
收稿日期: 2019-03-12
要求修回日期: 2019-10-15
网络出版日期: 2020-03-10
基金资助
海洋赤潮灾害立体监测技术与应用国家海洋局重点实验室开放研究基金(MATHAB201709)
海洋公益性行业科研专项(201305012)
版权
Variation characteristics of the evolution of Karenia mikimotoi bloom and environmental factors based on online monitoring buoy data*
Received date: 2019-03-12
Request revised date: 2019-10-15
Online published: 2020-03-10
Supported by
Foundation item: Key Laboratory of Integrated Monitoring and Applied Technology for Marine Harmful Algal Blooms, S.O.A.(MATHAB201709)
Public Science and Technology Research Funds Projects of Ocean(201305012)
Copyright
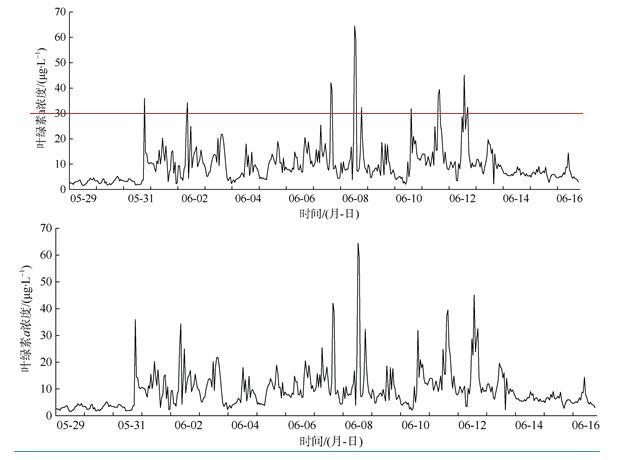
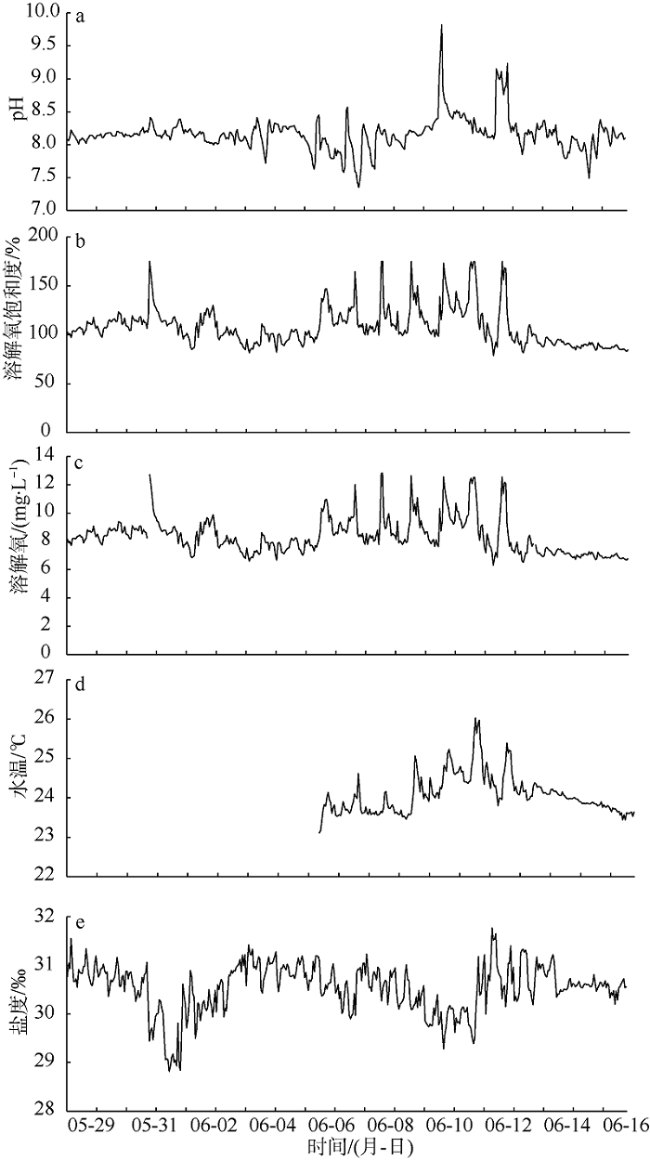
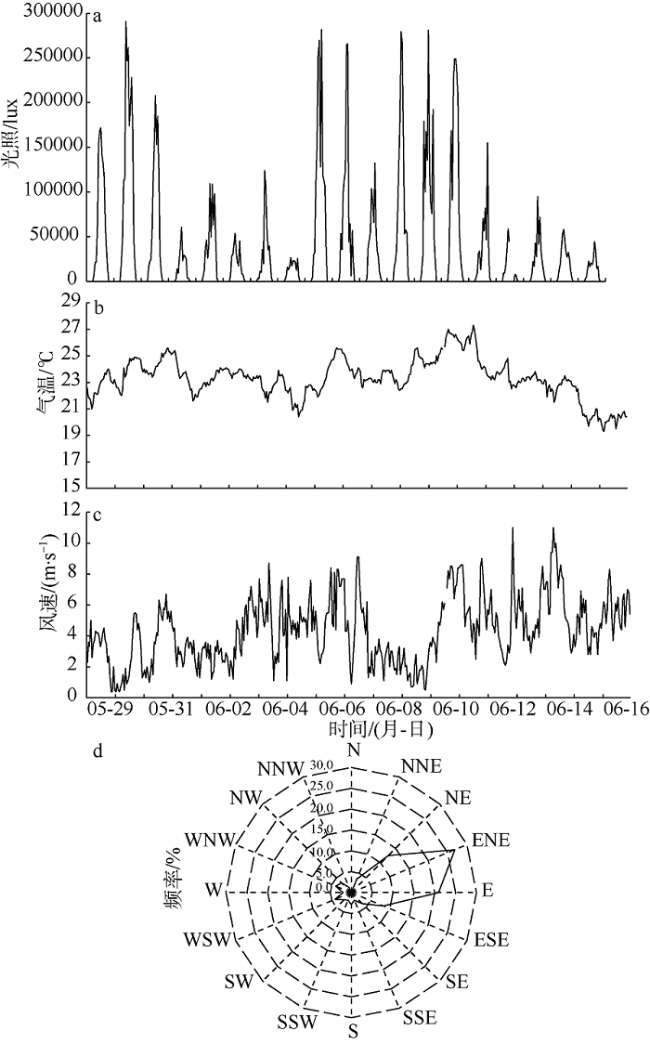
相较于船舶走航监测, 海洋水质浮标在线监测的优势在于能够获取目标海域长期、连续监测数据, 能更好地反映环境状况的动态变化。为了厘清米氏凯伦藻赤潮的发生、发展动态, 本文以2017年6月南麂附近海域米氏凯伦藻赤潮为例, 分析海洋水质浮标获取的连续监测数据, 探讨米氏凯伦藻赤潮过程叶绿素a和水环境因子动态变化特征及其与气象要素的关系。赤潮期间, 水温为22.8~26.0℃、盐度为28.8‰~31.8‰、气温为20.4~27.3℃, 该温盐范围均适宜米氏凯伦藻的细胞生长; 较高的光照强度能够支持较高的藻类细胞密度。Pearson相关性分析显示, 米氏凯伦藻细胞密度与叶绿素a浓度呈显著正相关; 溶解氧(DO)及其饱和度(DO%)、pH、水温、气温等环境因子与叶绿素a浓度呈极显著正相关, 盐度与叶绿素a浓度呈极显著负相关。向岸风生海流有利于藻种向近岸较高营养区域汇集, 为赤潮的爆发创造有利条件。赤潮过程中, 叶绿素a浓度、溶解氧饱和度、pH发生了协同变化, 据此特征可以开展赤潮短期预警。
赵聪蛟 , 刘希真 , 付声景 , 姚炜民 , 周燕 , 马骏 . 基于水质浮标在线监测的米氏凯伦藻赤潮过程及环境因子变化特征分析[J]. 热带海洋学报, 2020 , 39(2) : 88 -97 . DOI: 10.11978/2019027
Superior to traditional monitoring methods, the marine monitoring buoys are able to obtain long-term and real-time continuous marine environmental data to reflect the dynamic changes in marine environmental conditions. In order to clarify the occurrence and development of the red tide of Karenia mikimotoi, and to carry out integrated monitoring and early warning of red tide disasters, we analyzed the evolution of Karenia mikimotoi bloom and environmental factors based on online monitoring buoy data obtained from the coastal area of Nanji Island of Zhejiang Province in June 2017. K. mikimotoi bloom occurs in the sea water with temperature and salinity ranging from 22.8℃ to 26.0℃ and from 28.8 to 31.8, and when the air temperature is between 20.4℃ and 27.3℃, which are suitable for the cell growth of K. mikimotoi. Higher light intensity can support higher K. mikimotoi cell density. Pearson correlation analysis shows that the cell density of K. mikimotoi is significantly positively correlated with chlorophyll a (chl-a). Dissolved oxygen (DO), dissolved oxygen saturation (DO%), pH, water temperature, and air temperature are all significantly positively correlated with chl-a. Salinity is significantly negatively correlated with chl-a. Wind blowing ocean current may facilitate the accumulation of algae to the shore, which creates suitable conditions for the outbreak of red tides. During the red tide, chl-a, DO% and pH have synergistic changes, which may carry out red tide early-warning signs based on the online monitoring buoy data.
表1 2017年6月温州南麂附近海域米氏凯伦藻赤潮过程主要指标监测结果Tab. 1 Results of major monitoring indicators during the red tide in June 2017 |
| 时间 | 浮标监测数据 | 人工采样监测数据 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 叶绿素a浓度/(μg·L-1) | pH | DO% | DO /(mg·L-l) | 水温/℃ | 盐度/‰ | 光照/lx | 气温/℃ | 风速 /(m·s-1) | 风向/° | 最大细胞密度 /(×106个·L-1) | 面积/km2 | 采样点纬度 | 采样点经度 | |
| 6月2日 | 24.97 | 8.20 | 122.0 | 9.34 | 23.64 | 29.86 | 40630 | 23.3 | 3.3 | 47 | 9.10 | 1.0 | 27°28′38"N | 121°03′32"E |
| 6月6日 | 12.79 | 8.34 | 100.5 | 7.87 | 23.17 | 31.19 | 111880 | 23.3 | 5.5 | 110 | 6.30 | 25.0 | 27°27′13"N | 121°04′19"E |
| 6月9日 | 5.54 | 8.17 | 124.0 | 9.36 | 24.19 | 30.69 | 179800 | 24.8 | 2.8 | 88 | 0.33 | 0.0 | 27°27′13"N | 121°04′19"E |
| 6月12日 | 12.73 | 9.15 | 111.9 | 8.48 | 24.51 | 30.99 | 70510 | 23.7 | 2.5 | 285 | 2.20 | 5.0 | 27°28′40"N | 121°03′12"E |
| 6月13日 | 13.03 | 8.17 | 91.5 | 7.16 | 24.04 | 31.31 | 49930 | 23.4 | 2.9 | 23 | 1.70 | 3.5 | 27°28′40"N | 121°03′12"E |
| 6月15日 | 9.07 | 7.90 | 90.5 | 7.13 | 23.85 | 30.54 | 58150 | 20.6 | 4.1 | 59 | 0.46 | 0.0 | 27°28′40"N | 121°03′12"E |
表2 叶绿素a浓度与细胞密度及环境因子的Pearson相关性分析Tab. 2 Pearson's correlation analysis of chlorophyll a and cell density, and of chlorophyll a and environmental factors |
| 项目 | 叶绿素a浓度 | 细胞密度 | pH | DO% | DO | 水温 | 盐度 | 光照 | 气温 | 风速 | 风向 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 叶绿素a浓度 | 1.000 | 0.879* | 0.200** | 0.633** | 0.624** | 0.480** | -0.308** | 0.054 | 0.268** | 0.055 | -0.049 |
| p值 | p<0.001(N=451) | p=0.021(N=6) | p<0.001(N=451) | p=0.256(N=449) | p<0.001(N=450) | p=0.241(N=450) | p=0.300(N=450) | ||||
注: **表示极显著相关, p<0.01; *表示显著相关, p<0.05; N为样本数 |
| 1 |
陈宝红, 谢尔艺, 高亚辉 , 等, 2015. 米氏凯伦藻对海洋生物致毒作用的研究进展[J]. 福建水产, 37(3):241-250.
CHEN BAOHONG, XIE ERYI, GAO YAHUI, et al, 2015. Toxic effects of red tide caused by Karenia mikimotoi on marine organisms[J]. Journal of Fujian Fisheries, 37(3):241-250 (in Chinese with English abstract).
|
| 2 |
邓华, 管卫兵, 曹振轶 , 等, 2016. 2012年福建沿海大规模米氏凯伦藻赤潮暴发的水文气象原因探讨[J]. 海洋学研究, 34(4):28-38.
DENG HUA, GUAN WEIBING, CAO ZHENYI, et al, 2016. Analysis of hydrological and meteorological factors causing Karenia mikimotoi bloom in 2012 along Fujian coast[J]. Journal of Marine Sciences, 34(4):28-38 (in Chinese with English abstract).
|
| 3 |
丁光茂, 张树峰 , 2018. 2012年三沙湾米氏凯伦藻赤潮的生态特征及成因分析[J]. 海洋学报, 40(6):104-112.
DING GUANGMAO, ZHANG SHUFENG, 2018. Ecological characteristics and the causes of Karenia mikimotoi bloom in the Sansha bay in 2012[J]. Haiyang Xuebao, 40(6):104-112 (in Chinese with English abstract).
|
| 4 |
郭皓, 林凤翱, 刘永健 , 等, 2014. 近年来我国海域多发性赤潮生物种类以及赤潮风险指数分级预警方法[J]. 海洋环境科学, 33(1):94-98.
GUO HAO, LIN FENG’AO, LIU YONGJIAN, et al, 2014. High-incidenceHABs causative species in China coastal waters and the forewarning method based on the HABs Risk Index[J]. Marine Environmental Science, 33(1):94-98 (in Chinese with English abstract).
|
| 5 |
李涛, 刘胜, 黄良民 , 等, 2005. 大亚湾一次赤潮生消期间浮游植物群落变化研究[J]. 热带海洋学报, 24(3):18-24.
LI TAO, LIU SHENG, HUANG LIANGMIN, et al, 2005. Studies on phytoplankton community change at Daya bay during a red tide[J]. Journal of Tropical Oceanography, 24(3):18-24 (in Chinese with English abstract).
|
| 6 |
李天深, 李远强, 赖春苗 , 等, 2011. 廉洲湾赤潮自动监测结果与分析[J]. 中国环境监测, 27(4):32-35.
LI TIANSHEN, LI YUANQIANG, LAI CHUNMIAO, et al, 2011. Analyze red tide with automatic monitoring system of water quality in Lianzhou gulf[J]. Environmental Monitoring in China, 27(4):32-35 (in Chinese with English abstract).
|
| 7 |
李扬, 吕颂辉, 江天久 , 等, 2009. 南麂列岛海域原甲藻种群动态及其环境影响因子分析[J]. 水生生物学报, 33(2):236-245.
LI YANG, LV SONGHUI, JIANG TIANJIU, et al, 2009. Dynamics of prorocentrum population and its relation with environmental factors in Nanji Islands sea area[J]. Acta Hydrobiologica Sinica, 33(2):236-245 (in Chinese with English abstract).
|
| 8 |
刘桂英, 葛坤, 宋伦 , 等, 2015. 米氏凯伦藻的研究进展[J]. 海洋科学, 39(9):117-122.
LIU GUIYING, GE KUN, SONG LUN, et al, 2015. The research trend of Karenia mikimotoi[J]. Marine Sciences, 39(9):117-122 (in Chinese).
|
| 9 |
刘志国, 王金辉, 蔡芃 , 等, 2014. 米氏凯伦藻分布及其引发赤潮的发生规律研究[J]. 国土与自然资源研究, (1):38-41.
LIU ZHIGUO, WANG JINHUI, CAI PENG, et al, 2014. Distribution of Karenia mikimotoi and rules of its causing red tide[J]. Territory & Natural Resources Study, ( 1):38-41 (in Chinese with English abstract).
|
| 10 |
龙华, 杜琦 , 2005. 福建沿海米氏凯伦藻赤潮的初步研究[J]. 福建水产, (4):22-26.
LONG HUA, DU QI, 2005. Primary research on Karenia mikimotoi bloom in Fujian coast[J]. Journal of Fujian Fisheries, (4):22-26 (in Chinese with English abstract).
|
| 11 |
陆斗定,
LU DOUDING, GOBEL J, WANG CHUNSHENG, et al, 2000. Monitoring of harmful microalgae and nowcasting of red tides in Zhejiang coastal water[J]. Donghai Marine Science, 18(2):33-44 (in Chinese with English abstract).
|
| 12 |
吕颂辉, 黄凯旋 , 2007. 米氏凯伦藻在三种无机氮源的生长情况[J]. 生态环境, 16(5):1337-1341.
LV SONGHUI, HUANG KAIXUAN, 2007. The growth of Karenia mikimotoi Hansen in three different types of inorganic nitrogen sources[J]. Ecology and Environment, 16(5):1337-1341 (in Chinese with English abstract).
|
| 13 |
潘非斐, 吴楠 , 2017. 不同氮磷比对福建沿海米氏凯伦藻生长的影响[J]. 渔业研究, 39(4):287-294.
PAN FEIFEI, WU NAN, 2017. The effects of nitrogrn to phosphorus ratio on the growth of Karenia mikimotoi separated from Fujian coastal area[J]. Journal of Fisheries Research, 39(4):287-294 (in Chinese with English abstract).
|
| 14 |
庞勇, 聂瑞, 吕颂辉 , 2015. 珠江口双胞旋沟藻Cochlodinium geminatum赤潮生消过程的环境特征初步分析[J]. 生态环境学报, 24(2):286-293.
PANG YONG, NIE RUI, LV SONGHUI, 2015. Preliminary environmental analysis of the evolution of Cochlodinium geminatum bloom in the Pearl River estuary[J]. Ecology and Environmental Sciences, 24(2):286-293 (in Chinese with English abstract).
|
| 15 |
庞勇, 聂瑞, 吕颂辉 , 2016. 不同磷源对米氏凯伦藻生长和碱性磷酸酶活性的影响[J]. 海洋科学, 40(4):59-64.
PANG YONG, NIE RUI, LV SONGHUI, 2016. Effects of the different kinds of phosphorus sources on growth and Alkaline Phosphatase Activity (APA) of Karenia mikimotoi Hansen[J]. Marine Sciences, 40(4):59-64 (in Chinese with English abstract).
|
| 16 |
王汉奎, 黄良民, 黄小平 , 等, 2003. 珠江口海域条纹环沟藻赤潮的生消过程和环境特征[J]. 热带海洋学报, 22(5):55-62.
WANG HANKUI, HUANG LIANGMIN, HUANG XIAOPING, et al, 2003. A red tide caused by Gyrodinium instriatum and its environmental characters in Zhujiang river estuary[J]. Journal of Tropical Oceanography, 22(5):55-62 (in Chinese with English abstract).
|
| 17 |
王越, 沈盎绿, 赵世烨 , 等, 2015. pH对米氏凯伦藻(Karenia mikimotoi)种群生长、营养吸收及无机碳亲和力的影响[J]. 海洋环境科学, 34(4):488-493.
WANG YUE, SHEN ANGLV, ZHAO SHIYE, et al, 2015. Effects of pH limitation on population growth, nutrient uptake and photosynjournal physiological processes of Karenia mikimotoi[J]. Marine Environmental Science, 34(4):488-493 (in Chinese with English abstract).
|
| 18 |
文世勇, 刘希真, 王紫竹 , 等, 2018. 不同光照条件下米氏凯伦藻和东海原甲藻生长的温度生态幅[J]. 生态学报, 38(14):5187-5194.
WEN SHIYONG, LIU XIZHEN, WANG ZIZHU, et al, 2018. Temperature-ecological amplitude of Karenia mikimotoi and Prorocentrum donghaiense at different light intensities in laboratory culture[J]. Acta Ecologica Sinica, 38(14):5187-5194 (in Chinese with English abstract).
|
| 19 |
姚炜民, 潘晓东, 华丹丹 , 2007. 浙江海域米氏凯伦藻赤潮成因的初步研究[J]. 水利渔业, 27(6):57-58, 76.
YAO WEIMIN, PAN XIAODONG, HUA DANDAN, 2007. Preliminary studies on the formation cause of red tide by Karenia mikimotoi in the Zhejiang sea area[J]. Reservoir Fisheries, 27(6):57-58, 76 (in Chinese).
|
| 20 |
尹翠玲, 张秋丰, 邹涛 , 等, 2013. 渤海湾天津近岸海域夜光藻赤潮生消过程初探[J]. 海洋湖沼通报, ( 2):99-104.
YIN CUILING, ZHANG QIUFENG, ZOU TAO, et al, 2013. Analysis for Noctiluca Scintillans red tide in Bohai bay[J]. Transactions of Oceanology and Limnology, ( 2):99-104 (in Chinese with English abstract).
|
| 21 |
赵聪蛟, 孔梅, 孙笑笑 , 等, 2016 a. 浙江省海洋水质浮标在线监测系统构建及应用[J]. 海洋环境科学, 35(2):288-294.
ZHAO CONGJIAO, KONG MEI, SUN XIAOXIAO, et al, 2016a. Construction and application of the marine online monitoring buoy system in Zhejiang province[J]. Marine Environmental Science, 35(2):288-294 (in Chinese with English abstract).
|
| 22 |
赵聪蛟, 周燕, 姚炜民 , 等, 2016b. 一种基于浮标在线监测数据的赤潮短期预警方法及用途: 中国, 105403680A[P]. 2016-03-16 (in Chinese).
|
| 23 |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. GB 17378.3-2007 海洋监测规范第3部分: 样品采集、贮存与运输[S]. 北京: 中国标准出版社.
GENERAL ADMINISTRATION OF QUALITY SUPERVISION, INSPECTION AND QUARANTINE OF THE PEOPLE’S REPUBLIC OF CHINA, STANDARDIZATION ADMINISTRATION OF CHINA, 2008. GB 17378.3-2007 The specification for marine monitoring—Part 3: sample collection, storage and transportation[S]. Beijing: Standards Press of China (in Chinese).
|
| 24 |
周钦, 马增岭, 袁兴伟 , 等, 2017. 米氏凯伦藻生长对磷的响应及其吸收动力学研究[J]. 上海海洋大学学报, 26(4):546-553.
ZHOU QIN, MA ZENGLING, YUAN XINGWEI, et al, 2017. Studies on growth and phosphorous uptake kinetics of Karenia mikimotoi at the different phosphorous levels[J]. Journal of Shanghai Ocean University, 26(4):546-553 (in Chinese with English abstract).
|
| 25 |
朱德弟, 陆斗定, 王云峰 , 等, 2009. 2005年春初浙江近海的低温特征及其对大规模东海原甲藻赤潮发生的影响[J]. 海洋学报, 31(6):31-39.
ZHU DEDI, LU DOUDING, WANG YUNFENG, et al, 2009. The low temperature characteristics in Zhejiang coastal region in the early spring of 2005 and its influence on harmful algae bloom occurrence of Prorocentrum donghaiense[J]. Haiyang Xuebao, 31(6):31-39 (in Chinese with English abstract).
|
| 26 |
庄宏儒 , 2006. 水质自动监测系统在厦门同安湾赤潮短期预报中的应用[J], 海洋环境科学, 25(2):58-61.
ZHUANG HONGRU, 2006. Application of automatic monitoring system of water quality on short-term prediction of red tide in Tongan Bay, Xiamen[J]. Marine Environmental Science, 25(2):58-61 (in Chinese with English abstract).
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}