
南海北部海洋生态模型的参数分析及遗传算法优化
作者简介:舒婵(1994—), 女, 湖北省黄冈市人, 博士研究生, 主要从事海洋生态系统动力学研究。E-mail: shuchan@scsio.ac.cn
*感谢两位匿名审稿人提出宝贵的意见。
Editor: 殷波
收稿日期: 2019-06-06
要求修回日期: 2019-08-23
网络出版日期: 2020-03-10
基金资助
国家重点研发计划(2016YFC1401604)
国家自然科学基金(41576002、41730536、41890805)(41576002)
国家自然科学基金(41576002、41730536、41890805)(41730536)
国家自然科学基金(41576002、41730536、41890805)(41890805)
热带海洋环境国家重点实验室自主课题(LTOZZ1803)
版权
Parameter analysis and optimization using genetic algorithm in a marine ecosystem model of the northern South China Sea
Received date: 2019-06-06
Request revised date: 2019-08-23
Online published: 2020-03-10
Supported by
Foundation item: National Key Research and Development Program of China(2016YFC1401604)
Natural Science Foundation of China(41576002)
Natural Science Foundation of China(41730536)
Natural Science Foundation of China(41890805)
Foundation of State Key Laboratory of Tropical Oceanography(LTOZZ1803)
Copyright
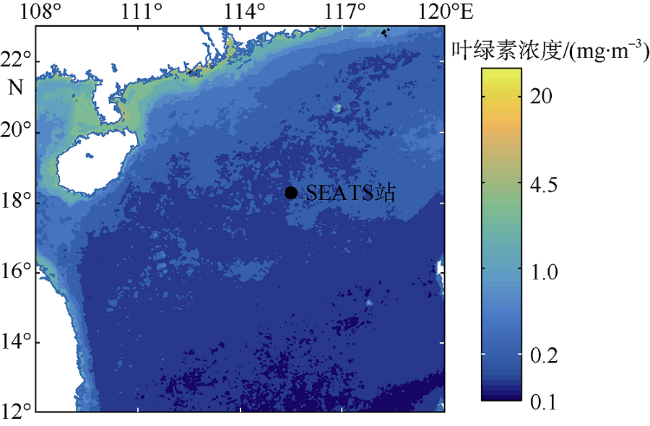
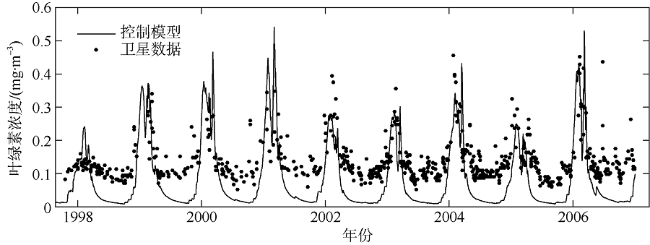
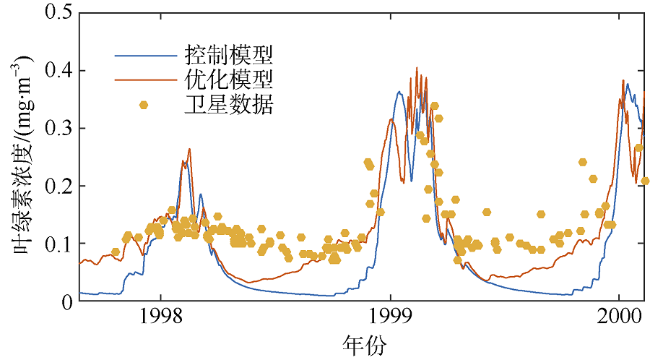
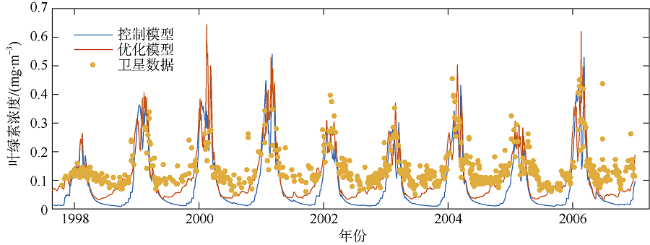
海洋生态系统动力学模型是研究海洋生态环境的重要手段。随着模型的发展, 生态参数取值不确定性增加, 对模型结果的影响逐渐增大, 因此模型参数优化显得尤为重要。本研究在南海北部应用一维物理-生态耦合模型, 通过对模型生态参数进行敏感性分析, 获取关键生态参数, 利用遗传算法对参数进行优化。结果表明, 模型中的敏感参数主要集中于浮游植物生长和浮游动物生长、摄食和死亡以及碎屑沉降等过程。针对以上参数利用遗传算法优化, 发现仅加入表层卫星数据, 模型表层和垂向模拟误差分别降低27.80%和21.40%; 加入垂向观测数据, 表层和垂向模拟误差分别降低14.90%和32.70%。遗传算法应用于海洋生态模型的关键参数优化研究, 所获取的参数对模型有明显的改善效果, 提高了耦合模型对生态系统的模拟精度, 为参数优化在三维模型中的应用提供了依据。
舒婵 , 耿兵绪 , 房巍巍 , 修鹏 . 南海北部海洋生态模型的参数分析及遗传算法优化[J]. 热带海洋学报, 2020 , 39(2) : 98 -106 . DOI: 10.11978/2019054
Marine ecosystem dynamics model is an important means to study marine ecological environment. As the model complexity increases, the number and uncertainty of biological parameters increase, which has a great impact on model results; therefore, optimization of model parameters is particularly important. In this paper, a one-dimensional physical-biological model is applied to the northern South China Sea, and the key biological parameters obtained through sensitivity analysis are optimized by using genetic algorithm. The results show that the sensitive parameters in the model are related to phytoplankton growth, zooplankton growth, feeding and death, and detritus sinking. Based on the genetic algorithm optimization of the above-mentioned parameters, we find that the surface and vertical simulation errors of the model are reduced by 27.80% and 21.40%, respectively, by using only surface satellite data; the surface and vertical simulation errors are reduced by 14.90% and 32.70%, respectively, by adding observed profile data. The success of applying genetic algorithm in the one-dimensional model provides the basis for its further application in three-dimensional marine ecosystem models.
表1 生态参数取值以及敏感度Tab. 1 Values and sensitivities of biological parameters |
| 参数 | 描述/(单位) | 取值 | 敏感度 |
|---|---|---|---|
| akz2 | Z2捕食半饱和常数/(mmol_N·m-3) | 0.25 | -16.55 |
| reg2 | Z2铵盐排出速率/d-1 | 0.05 | -2.00 |
| Chl2cs2_m | S2叶绿素同碳比值最大值/(mg_Chl·mg_C-1) | 6.50×10-2 | -1.59 |
| beta2 | Z2最大捕食速率/d-1 | 0.53 | 1.11 |
| bgamma2 | Z2捕食效率 | 0.75 | 1.11 |
| bgamma1 | Z1捕食效率 | 0.75 | 1.04 |
| beta1 | Z1最大捕食速率/d-1 | 0.80 | 1.01 |
| Chl2cs1_m | S1叶绿素同碳比值最大值/(mg_Chl mg_C-1) | 6.50×10-2 | 0.99 |
| wsd | 碎屑下沉速率/(m·d-1) | 10.00 | -0.84 |
| aknh4s1 | S1吸收铵盐半饱和常数/(mmol_N·m-3) | 0.05 | 0.71 |
| akz1 | Z1捕食半饱和常数/(mmol_N·m-3) | 0.50 | 0.64 |
| reg1 | Z1铵盐排出速率/d-1 | 0.10 | 0.49 |
| bgamma5 | 碎屑衰减速率/d-1 | 0.05 | 0.45 |
| gmaxs1 | S1最大特征生长速率/d-1 | 2.00 | 0.37 |
| gmaxs2 | S2最大特征生长速率/d-1 | 3.00 | 0.27 |
| amaxs1 | S1 P-I曲线初始斜率/(m2·W-1·d-1) | 2.50×10-2 | -0.26 |
| akox | 氧化作用半饱和常数/(mmol_O·m-3) | 30.00 | 0.23 |
| ak2 | 浮游植物特征光衰减速率/(m2·mmol-1) | 0.03 | 0.22 |
| akno3s1 | S1吸收硝酸盐半饱和常数/(mmol_N·m-3) | 1.00 | -0.22 |
| akno3s2 | S2吸收硝酸盐半饱和常数/(mmol_N·m-3) | 2.00 | 0.21 |
| pis2 | S2铵盐抑制系数 | 3.00 | 0.14 |
| bgamma4 | S2死亡率/d-1 | 0.01 | 0.18 |
| wsp | S2下沉速率/(m·d-1) | 1.00 | 0.13 |
| bgamma0 | Z2特征死亡率/d-1 | 0.08 | -0.13 |
| aksio4s2 | S2吸收硅酸盐半饱和常数/(mmol·m-3) | 4.00 | 0.13 |
| parsats1/s2 | S1/S2光合有效辐射饱和初始参数/(W·m-2) | 40.00 | 0.13 |
| akpo4s1/s2 | S1/S2吸收硝酸盐半饱和常数/(mmol·m-3) | 0.08 | 0.13 |
| akco2s1/s2 | S1/S2吸收二氧化碳半饱和常数/(mmol·m-3) | 500.00 | 0.13 |
| wsdsi | 碎屑硅下沉速率/(m·d-1) | 60.00 | 0.13 |
| bgamma7 | 硝化作用速率/d-1 | 0.08 | 0.10 |
| pis1 | S1铵盐抑制系数 | 1.50 | 0.08 |
| bgamma6 | 浮游植物凝聚速率/d-1 | 0.05 | 0.05 |
| bgamma3 | S1死亡率/d-1 | 0.01 | 0.02 |
注: CoSiNE模型的生态参数取值参考前人研究(Fasham, 1995; Evans et al, 1985; Chai et al, 2002; Kishi et al, 2007) |
表2 关键参数及其取值范围Tab. 2 Key parameters and their ranges |
| 参数 | bgamma2 | akz2 | beta2 | bgamma1 | Chl2cs1_m | wsd | reg2 | amaxs1 | gmaxs1 |
|---|---|---|---|---|---|---|---|---|---|
| r2 | 0.11 | 0.09 | 0.06 | 0.04 | 0.03 | 0.02 | 0.02 | 0.01 | 0.01 |
| 下限 | 0.37 | 0.12 | 0.25 | 0.37 | 0.03 | 5.00 | 0.02 | 0.01 | 1.00 |
| 上限 | 1.00 | 0.50 | 1.00 | 1.00 | 0.08 | 20.00 | 0.10 | 0.05 | 4.00 |
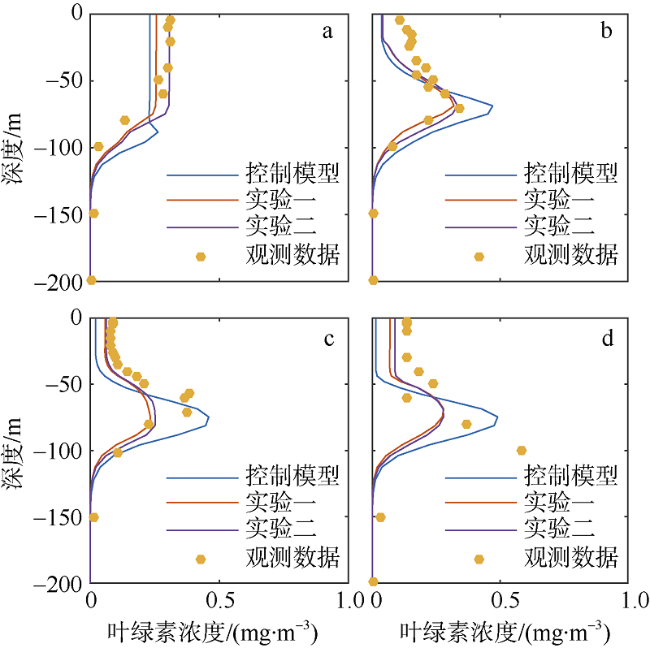
图5 垂向优化结果对比图a. 冬季(2003年1月20日); b. 春季(2000年5月23日); c. 夏季(2000年7月25日); d. 秋季(2000年10月18日) Fig. 5 Comparison of vertical optimization chlorophyll concentration. (a) winter; (b) spring; (c) summer; and (d) autumn |
表3 优化参数取值以及变化率Tab. 3 Optimization parameter values and rate of change |
| 参数 | bgamma2 | akz2 | beta2 | bgamma1 | Chl2cs1_m | wsd | reg2 | amaxs1 | gmaxs1 |
|---|---|---|---|---|---|---|---|---|---|
| 控制模型取值 | 0.75 | 0.25 | 0.53 | 0.75 | 6.50×10-2 | 10.00 | 0.05 | 2.50×10-2 | 2.00 |
| 实验一值 | 0.85 | 0.30 | 0.59 | 0.74 | 0.05 | 10.95 | 0.06 | 2.20×10-2 | 3.28 |
| 变化率 | 1.14 | 1.22 | 1.12 | 0.99 | 0.75 | 1.09 | 1.20 | 0.88 | 1.64 |
| 实验二值 | 0.86 | 0.28 | 0.49 | 0.78 | 0.05 | 12.07 | 0.06 | 2.40×10-2 | 3.44 |
| 变化率 | 1.15 | 1.14 | 0.93 | 1.04 | 0.84 | 1.21 | 1.20 | 0.96 | 1.72 |
| 1 |
高会旺, 孙文心, 翟雪梅 , 1999. 水层生态系统动力学模式参数的敏感性分析[J]. 青岛海洋大学学报(自然科学版) 398-404
GAO HUIWANG, SUN WENXIN, ZHAI XUEMEI, 1999. Sensitive analysis of the parameters of a pelagic ecosystem dynamic model[J]. Journal of Ocean University of Qingdao: 398-404 (in Chinese).
|
| 2 |
黄邦钦, 柳欣 , 2015. 边缘海浮游生态系统对生物泵的调控作用[J]. 地球科学进展, 30(3):385-395.
HUANG BANGQIN, LIU XIN, 2015. Review on planktonic ecosystem and its control on biological pump in the marginal seas[J]. Advances in Earth Science, 30(3):385-395 (in Chinese with English abstract).
|
| 3 |
尚毅威, 肖武鹏, 柳欣 , 等, 2018. 南海SEATS站超微型浮游植物类群及其碳生物量和叶绿素a浓度比值的变化[J]. 厦门大学学报(自然科学版), 57: 811-818.
SHANG YIWEI, XIAO WUPENG, LIU XIN, et al, 2018. Variations of pico-phytoplankton and carbon to chloropjyll-a ratios in the South China Sea at the SEATS staition[J]. Journal of Xiamen University, 57:811-818 (in Chinese).
|
| 4 |
张武昌, 张翠霞, 肖天 , 2009. 海洋浮游生态系统中小型浮游动物的生态功能[J]. 地球科学进展, 24:1195-1201.
ZhANG WUCHANG, ZHANG CUIXIA, XIAO TIAN, 2009. Role of Microzooplankton in Marine Planktonic Ecosystem[J]. Advances in Earth Science, 24:1195-1201 (in Chinese with English abstract).
|
| 5 |
赵亮 , 2002. 渤海浮游植物生态动力学模型研究[D] 青岛:青岛海洋大学.
ZHAO LIANG, 2002. A modeling study of the phytoplankton dynamic in the Bohai Sea[D] . Qingdao: Ocean University of Qingdao (in Chinese with English abstract).
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}