
大亚湾夏季浮游植物群落结构及对淡澳河输入的响应特征*
|
张立明(1994—), 男, 广东省梅州市人, 硕士研究生, 从事海洋生物地球化学研究。E-mail: |
Copy editor: 殷波
收稿日期: 2020-01-21
要求修回日期: 2020-03-30
网络出版日期: 2020-04-01
基金资助
国家自然科学基金项目(31971432)
国家自然科学基金项目(41506161)
南方海洋科学与工程广东省实验室(广州)项目(GML2019ZD0405)
广东省科技计划项目(2017B0303014052)
广东省海洋经济促进项目基金(GDOE2019A32)
版权
Characteristics of the phytoplankton community and its response to Dan’ao River input in Daya Bay in summer*
Copy editor: YIN Bo
Received date: 2020-01-21
Request revised date: 2020-03-30
Online published: 2020-04-01
Supported by
National Natural Science Foundation of China(31971432)
National Natural Science Foundation of China(41506161)
Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0405)
Science and Technology Planning Project of Guangdong Province, China(2017B0303014052)
Guangdong marine economy promotion projects Fund(GDOE2019A32)
Copyright
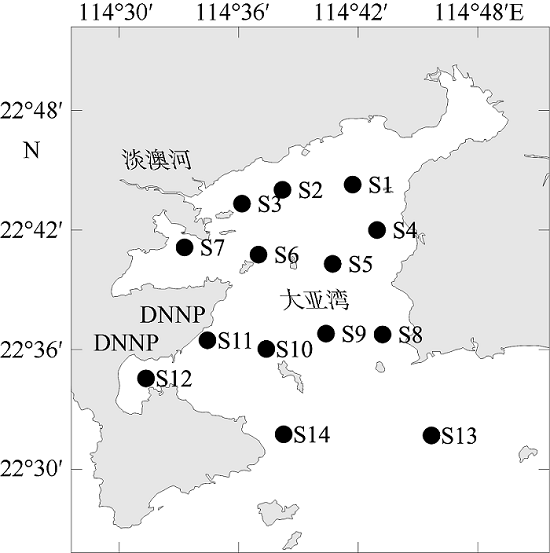
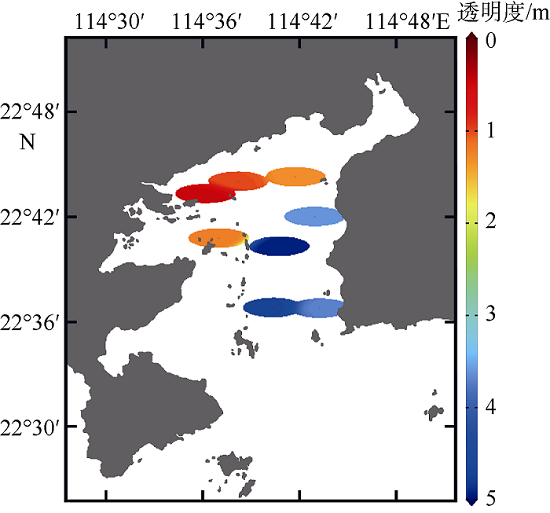
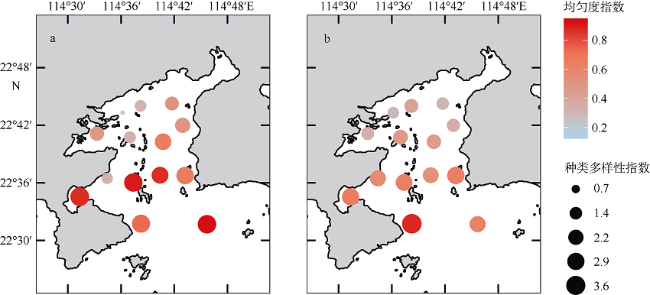
浮游植物是水生生态系统的基础生产者, 其群落结构直接影响到生态系统的健康和安全。河流输入是人类活动影响大亚湾水体环境最重要的途径之一, 淡水输入改变了水体温度、盐度、浊度和营养盐等环境因子, 对浮游植物群落结构产生影响。文章调查研究了2015年河流输入最强的夏季丰水期大亚湾的水体环境因子和浮游植物群落结构, 分析了在较强河流输入影响下浮游植物群落结构的动态变化及其对环境因子的响应。结果发现, 夏季大亚湾淡澳河的输入使湾顶淡澳河口区域形成层化的低盐、高温、低透明度、高营养盐的水体, 湾中部表层水体则受一定强度河流羽流影响, 而湾口和湾中部底层水体主要受外海水影响。淡澳河淡水输入是夏季大亚湾外源性氮、磷营养盐的主要来源, 而硅酸盐除河流输入外, 外海水也输入较多的营养盐使得底层水体硅酸盐浓度较高。夏季大亚湾水体营养比例失衡较严重, 溶解无机磷是限制浮游植物生长的重要因子。硅藻是大亚湾夏季浮游植物的优势类群, 调查发现3种优势种[极小海链藻(Thalassiosira minima)、中肋骨条藻(Skeletonema costatum)和圆海链藻(Thalassiosira rotula)]均为硅藻。通过聚类分析, 可将大亚湾夏季浮游植物群落主要分为3种类型, 分别为: 浮游植物丰度较大的极小海链藻藻华暴发的群落, 位于淡澳河口, 受河流输入影响明显; 中肋骨条藻占据优势的群落, 分布在受一定强度的河流及其羽流影响的湾顶和湾中部区域; 浮游植物丰度较低的群落, 无明显优势种, 主要分布在湾口海水影响区域。淡澳河口的水体环境有利于小型链状硅藻极小海链藻的快速繁殖并暴发了藻华, 藻华发生时的海水环境条件为: 温度30~31°C, 盐度17‰~31‰, 水体透明度0.45~1.2m。硅藻对不同营养盐利用的差异以及随后的生物碎屑和颗粒沉降过程导致藻华发生区域Si∶N值略降低, N∶P值显著升高。河流输入影响下, 单一物种大量生长使得浮游植物群落种类组成丰度分布极不均匀, 从而导致淡澳河口浮游植物群落的种类多样性和均匀度指数降低, 种类多样性和均匀度指数均从淡澳河口向湾口逐渐增大。
张立明 , 谭烨辉 , 李佳俊 , 黄小平 , 刘甲星 . 大亚湾夏季浮游植物群落结构及对淡澳河输入的响应特征*[J]. 热带海洋学报, 2020 , 39(5) : 43 -54 . DOI: 10.11978/2020012
Phytoplankton are the primary producer of aquatic ecosystem, and their community structure directly affects the health and safety of aquatic ecosystem. River input is one of the most important impacts reflecting the human activities on Daya Bay waters, which influences the phytoplankton community structure by changing environmental factors such as temperature, salinity, turbidity, nutrients, and so on. We investigated water environmental factors and phytoplankton community structure in Daya Bay in the summer of 2015 when large amount of river input happened. The dynamic change of phytoplankton community structure under the influence of the strong river input and its response to environmental factors were analyzed. The results showed that stratified waters with low salinity, high temperature, declined transparency, and enhanced nutrient concentrations formed at the Dan’ao River Mouth due to river input, while the waters in the middle and opening of Daya Bay were greatly affected by seawater intrusion. Freshwater input was the main external source of nitrogen and phosphorus nutrients in Daya Bay, and both freshwater input and invasive seawater were both important source of silicate. Nutrient structure was highly imbalanced in Daya Bay in summer, and dissolved inorganic phosphorus was the important limited factor of the growth of phytoplankton. Diatoms were the dominant group of phytoplankton in Daya Bay in summer 2015, and three dominant species included Thalassiosira minima, Skeletonema costatum and Thalassiosira rotula were identified. Cluster analysis showed that phytoplankton community in Daya Bay in summer 2015 could be mainly divided into three types. Type Ⅰ: Algae assemblage occurred at the Dan’ao River Mouth with high abundance of phytoplankton and bloom of Thalassiosira minima under influence of freshwater discharge; Type Ⅱ: algae assemblage dominated by Skeletonema costatum observed at the top and middle Daya Bay under subordinate influence of river plume; Type Ⅲ: algae assemblage with low phytoplankton abundance and no obvious dominant species mainly occurred in the opening of Daya Bay under the influence of sea water. The water environments at the Dan’ao River Mouth were beneficial to the growth and bloom of tiny chain-formed Thalassiosira minima, and the bloom occurred in the environmental gradient of temperature 30 ~ 31 °C, salinity 17 ‰ ~ 31 ‰ and water transparency 0.45 ~ 1.2 m. The difference in the utilization of different nutrients by diatoms and the subsequent settlement of biogenic detritus and particles resulted in a slight decrease of Si∶N ratio and a significant increase of N∶P ratio in the area where algae bloomed. The distribution of phytoplankton abundance was extremely uneven at the Dan’ao River Mouth due to the bloom of single species under the influence of freshwater discharge, which led to the decrease of Shannon-Wiener index and Pielou’s evenness index. The two indexes both increased from the Dan’ao River Mouth to the opening of Daya Bay.
Key words: Daya Bay; phytoplankton; river input; community structure; nutrients structure
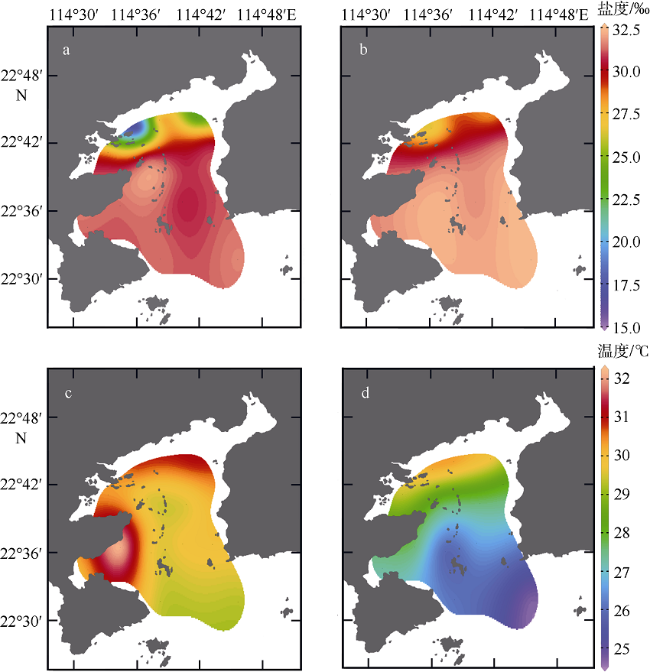
图2 大亚湾夏季海水温盐分布特征a. 表层盐度; b. 底层盐度; c. 表层温度; d. 底层温度。审图号为GS(2019)4343 Fig. 2 Distributions of salinity and water temperature in Daya Bay in summer 2015. (a) salinity of surface water; (b) salinity of bottom water; (c) temperature of surface water, and (d) temperature of bottom water |
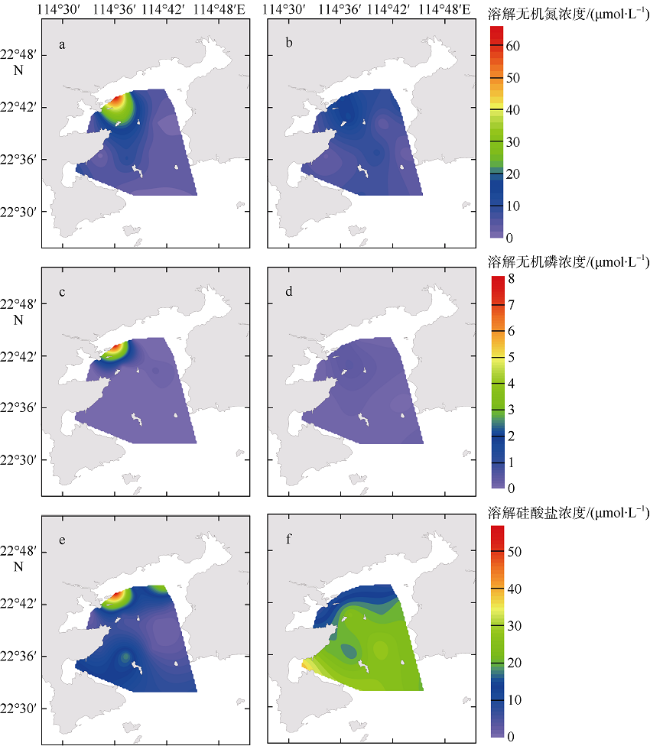
图4 大亚湾夏季海水营养盐浓度分布a. 表层溶解无机氮; b. 底层溶解无机氮; c. 表层溶解无机氮; d. 底层溶解无机氮; e. 表层硅酸盐; f. 底层硅酸盐。审图号为GS(2019)4343 Fig. 4 Distribution of nutrients in Daya Bay in summer 2015. (a) DIN in surface water; (b) DIN in bottom water; (c) DIP in surface water; (d) DIP in bottom water; (e) DSi in surface water, and (f) DSi in bottom water |
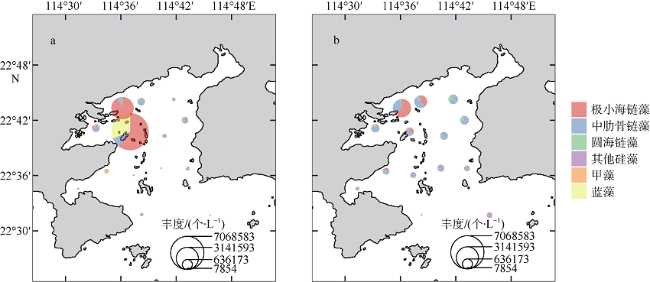
图5 大亚湾夏季浮游植物丰度及群落组成分布a. 表层水体; b. 底层水体。饼图大小表示各站位浮游植物丰度大小, 组成表示不同浮游植物群或物种的贡献比例。审图号为GS(2019)4343 Fig. 5 Distribution of phytoplankton abundance and composition in Daya Bay. (a) surface water; (b) bottom water. Pie size indicates the total abundance at each site, and composition indicates abundance contributions of different phytoplankton groups or species |
表1 2015年夏季大亚湾优势种及优势度Tab. 1 Dominant species and corresponding dominance in summer 2015 |
| 优势种 | 学名 | 优势度 |
|---|---|---|
| 极小海链藻 | Thalassiosira minima | 0.42 |
| 中肋骨条藻 | Skeletonema costatum | 0.25 |
| 圆海链藻 | Thalassiosira rotula | 0.04 |
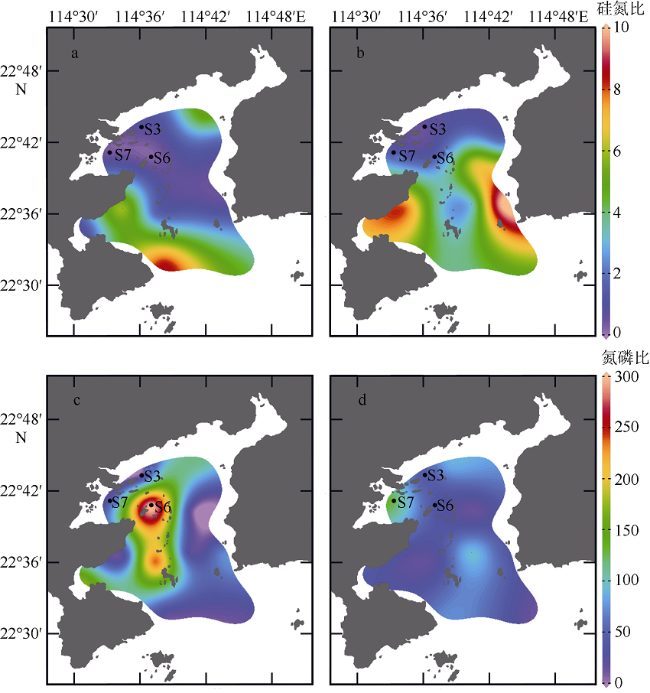
图6 大亚湾夏季营养盐结构及分布a. 表层水体Si:N; b. 底层水体Si:N; c. 表层水体N:P; d. 底层水体N:P。审图号为GS(2019)4343 Fig. 6 Distribution of different nutrients structure in Daya Bay in summer 2015. (a) Si:N ratio of surface water; (b) Si:N ratio of bottom water; (c) N:P ratio of surface water, and (d) N:P ratio of bottom water |
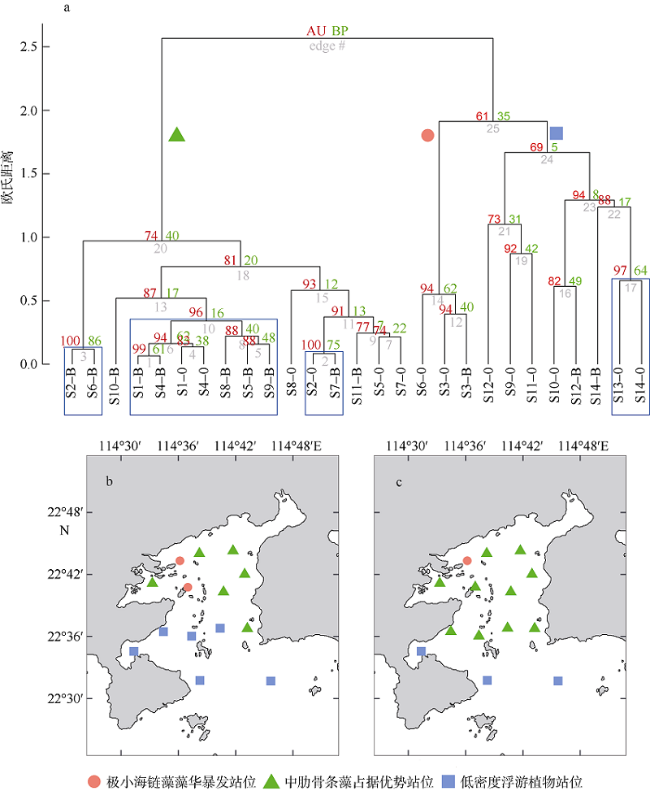
图7 大亚湾夏季浮游植物群落类型及其分布a. 带p值(单位: %)的聚类分析, 结果表明夏季大亚湾浮游植物群落可主要分为3类, 分别用绿色三角形、红色圆形和蓝色方形表示。站位名-S表示该站位表层水体, 站位名-B表示该站位底层水体。聚类树分支处左侧红色数值为AU (approximately unbiased)值, 右侧绿色数值为BP (bootstrap probability)值, 中间灰色数值表示聚类层次, 蓝色矩形框表示AU≥95%的集合; b. 不同类型浮游植物群落在表层水体分布; c. 不同类型浮游植物群落在底层水体分布。审图号为GS(2019)4343 Fig. 7 Distribution of different types of phytoplankton composition in Daya Bay in summer 2015. (a) cluster analysis with p values (units: %) showing phytoplankton community could mainly be classified into 3 types, which are represented by green triangle, red circular and blue square. Values at branches include AU p-values (left) and BP values (right), and gray values in the middle represent the clustering hierarchy. Clusters with AU ≥ 95 % are indicated by the rectangles; (b) surface water, and (c) bottom water |
| [1] |
白美娜, 江涛, 陈飞羽, 等, 2019. 大亚湾大鹏澳牡蛎养殖临近海域自养微微型浮游生物种群分布特征[J]. 海洋与湖沼, 50(1):129-138.
|
| [2] |
毕永红, 邓中洋, 胡征宇, 等, 2005. 发状念珠藻对盐胁迫的响应[J]. 水生生物学报, 29(2):125-129.
|
| [3] |
郝彦菊, 唐丹玲, 2010. 大亚湾浮游植物群落结构变化及其对水温上升的响应[J]. 生态环境学报, 19(8):1794-1800.
|
| [4] |
黄小平, 黄良民, 2002. 河口最大浑浊带浮游植物生态动力过程研究进展[J]. 生态学报, 22(9):1527-1533.
|
| [5] |
黄小平, 黄良民, 宋金明, 等, 2019. 营养物质对海湾生态环境影响的过程与机理[M]. 北京: 科学出版社: 1-20(in Chinese).
|
| [6] |
李丽, 江涛, 吕颂辉, 2013. 大亚湾海域夏、秋季分粒级叶绿素a分布特征[J]. 海洋环境科学, 32(2):185-189.
|
| [7] |
刘胜, 黄晖, 黄良民, 等, 2006. 大亚湾核电站对海湾浮游植物群落的生态效应[J]. 海洋环境科学, 25(2):9-12, 25.
|
| [8] |
唐森铭, 严岩, 陈彬, 2013. 春夏季大亚湾核电厂温排水对海洋浮游植物群落结构的影响[J]. 应用海洋学学报, 32(3):373-382.
|
| [9] |
王友绍, 王肇鼎, 黄良民, 2004. 近20年来大亚湾生态环境的变化及其发展趋势[J]. 热带海洋学报, 23(5):85-95.
|
| [10] |
谢福武, 宋星宇, 谭烨辉, 等, 2019. 模拟升温和营养盐加富对大亚湾浮游生物群落代谢的影响[J]. 热带海洋学报, 38(2):48-57.
|
| [11] |
杨熙, 谭烨辉, 2019. 夏季外海水入侵对大亚湾浮游植物群落结构的影响[J]. 海洋科学, 43(7):96-105.
|
| [12] |
杨雪, 王朝晖, 马长江, 等, 2016. 大亚湾微表层浮游植物对无机氮磷的响应[J]. 生态科学, 35(1):34-40.
|
| [13] |
张玉宇, 吕颂辉, 齐雨藻, 2008. 2003—2004年大亚湾澳头养殖区水域浮游植物群落结构及数量变动特征[J]. 海洋环境科学, 27(2):131-134.
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}