
一株群体感应抑制活性海洋放线菌的筛选与鉴定 *
|
李艳群(1992—), 女, 广东省英德市人, 硕士研究生, 研究方向为海洋生物活性物质及其化学生态学。email: |
Copy editor: 姚衍桃
收稿日期: 2020-01-20
要求修回日期: 2020-04-30
网络出版日期: 2020-05-19
基金资助
中国科学院战略性先导科技专项(A类)(XDA13020300)
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0401)
广东省重点领域研发计划(2020B1111030004)
版权
Screening and identification of a quorum sensing inhibitory actinomycetes derived from marine sediments
Received date: 2020-01-20
Request revised date: 2020-04-30
Online published: 2020-05-19
Supported by
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA13020300)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory(Guangzhou)(GML2019ZD0401)
Key Area Research and Development Program of Guangdong Province(2020B1111030004)
Copyright
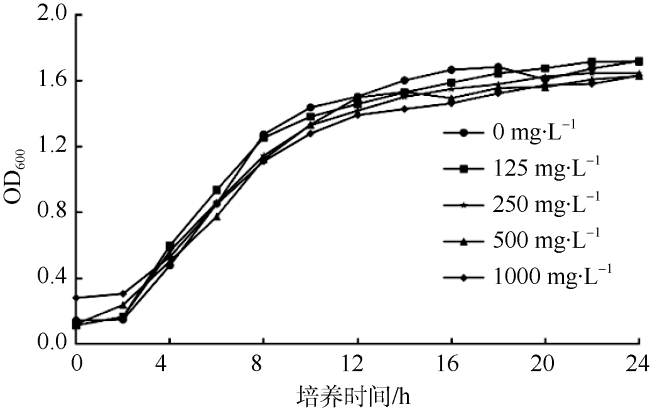
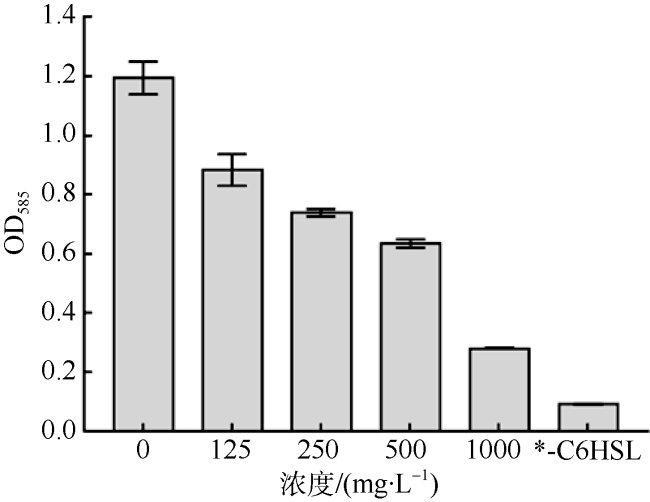
群体感应抑制剂(Quorum Sensing Inhibitors, QSIs)可以通过干扰群体感应(Quorum Sensing, QS)而有效降低病菌的感染性和毒力, 并且不会胁迫致病菌使其产生耐药性, 是具有前景的抗生素替代品。海洋微生物是新型QSIs的潜在来源。本研究利用紫色杆菌(Chromobacterium violaceum 026, CV026)模型评价QS抑制活性, 在来源于南海中西部和印度洋深海沉积物样品的放线菌中筛选QS抑制活性菌株, 并且根据形态学特征和16S rRNA基因序列鉴定活性菌株。通过筛选获得了菌株SCSIO 53717, 其提取物在CV026模型中能够显著降低指示菌株的紫色菌素产量, 并且在0~1000mg·mL-1的有效浓度范围内不影响紫色杆菌的生长。菌株SCSIO 53717被鉴定为Kocuria属, 是首次报道的具有QS抑制活性的Kocuria属菌株, 因而具有进一步深入研究的价值。
李艳群 , 陈柔雯 , 林宗豪 , 田新朋 , 尹浩 . 一株群体感应抑制活性海洋放线菌的筛选与鉴定 *[J]. 热带海洋学报, 2021 , 40(1) : 75 -81 . DOI: 10.11978/2020011
Quorum Sensing Inhibitors (QSIs) are promising alternatives to antibiotics, which can reduce the infectivity and virulence of pathogenic bacteria by interruption of quorum sensing (QS) pathways. The strategy aimed at disarming rather than at eradicating bacterial pathogens, and imposed no strong selective pressure on bacteria that can lead to the development of resistance mechanisms. Marine microorganism is one of the most potential sources of novel QSIs. In the present study, actinomycetes strains, isolated from the deep-sea sediment samples obtained from the South China Sea and Indian Ocean, were tested by Chromobacterium violaceum 026 (CV026) model, for screening for QS inhibitory strains. Strain SCSIO 53717 showed inhibitory activity against quorum sensing of CV026. The extract of SCSIO 53717 significantly reduced the production of CV026 violacein, and the extract of the strain showed no inhibitory effects on the growth of CV026 strain within the concentration range from 0 to 1000 mg·mL-1. This strain was assigned to the genus Kocuria based on morphological characteristics and 16S rRNA gene sequence analysis. To the best of our knowledge, this is the first time that Kocuria is reported to be QS inhibitory active. This result indicates that strain SCSIO 53717 is worthy further studying.
图1 SCSIO 53717提取物抑制紫色菌素的产生图中数字1、2、3、4、5依次表示浓度为25、50、100、150、200mg·mL-1的SCSIO 53717提取物(5μL); 6为阳性对照(加入5μL呋喃酮C30); 7为阴性对照(加入5μL甲醇溶液) Fig. 1 Inhibition of violacein production by SCSIO 53717 extracts. The numbers 1, 2, 3, 4, and 5 in the figure indicate the SCSIO 53717 extract (5μL) with a concentration of 25, 50, 100, 150, and 200 mg·mL-1, respectively; the number 6 is the positive control (adding 5μL of furanone C30); and the number 7 is the negative control (adding 5μL of methanol solution) |
图6 基于菌株SCSIO 53717及其近缘属种以16S rRNA基因序列构建的系统进化树线段0.005代 Fig. 6 Neighbor-Joining tree constructed showing the phylogenetic relationships among SCSIO 53717 strain and other related strains based on 16S rRNA gene sequence. Scale bar: 0.005 nucleotide substitution per 200 nucleotides of 16S rRNA gene sequence. Numerals on branches are the supporting percentage by 1000 replicates |
| [1] |
姚蓉, 马明, 付晖, 等, 2017. 海洋放线菌Kocuria sp.次级代谢产物的研究[J]. 中国海洋药物, 36(3):11-17.
|
| [2] |
尹守亮, 2011. 群体感应抑制活性海洋微生物的筛选与抑制因子活性评价[D]. 青岛: 中国海洋大学.
|
| [3] |
张炼辉, 2019. 微生物群体感应系统的研究进展[J]. 华南农业大学学报, 40(5):50-58.
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
/
| 〈 |
|
〉 |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}