
三亚湾肥胖软箭虫成体与幼体现场摄食差异研究
|
王崚力(1992—), 女, 四川省乐山市人, 博士研究生, 主要从事浮游动物摄食生态学研究。E-mail: wanglingli@scsio.ac.cn |
收稿日期: 2019-09-20
要求修回日期: 2019-12-06
网络出版日期: 2020-05-19
基金资助
中国科学院战略性先导科技专项(XDA13020100)
国家重点研发计划资助(2016YFC0502800)
国家重点研发计划资助(2017YFC0506302)
广东省省级科技计划项目(2017B0303014052)
国家自然科学基金项目(41806188)
中国科学院南海生态环境工程创新研究院自主部署项目(ISEE2018PY01)
版权
In situ feeding differences between adults and juveniles of chaetognath (Flaccisagitta enflata) in Sanya Bay
Received date: 2019-09-20
Request revised date: 2019-12-06
Online published: 2020-05-19
Supported by
The Strategic Priority Research Program of the Chinese Academy of Sciences(XDA13020100)
National Key Research and Development Project of China(2016YFC0502800)
National Key Research and Development Project of China(2017YFC0506302)
Science and Technology Planning Project of Guangdong Province, China(2017B0303014052)
National Natural Science Foundation of China(41806188)
Innovation Academy of South China Sea Ecology and Environmental Engineering, Chinese Academy of Sciences(ISEE2018PY01)
Copyright
毛颚类是热带近岸海域中上层鱼类的食物来源, 同时又是重要的肉食性浮游动物, 数量丰富, 然而其维持种群结构的生存策略尚不清晰。本文运用分子生物学技术, 分析了三亚湾海域毛颚类优势种肥胖软箭虫(Flaccisagitta enflata)成体与幼体的肠道食物组成, 以期从食物资源利用的角度揭示其维持种群结构的营养策略。研究结果显示, 幼体和成体摄食的浮游生物种类分别为21种和19种, 共同的食物类群有桡足类、小型水母类、硅藻和多毛类。成体与幼体食物偏好差异显著, 成体主要的食物来源是小型水母类(59%), 而幼体主要的食物来源是桡足类(60%); 幼体营养生态位(5.16)高于成体(2.89), 且二者食物重叠度低(0.21), 表明成体与幼体食物分化明显。研究结果揭示, 即使成体与幼体大量共存, 它们也可以通过摄食分化避免食物资源竞争, 这种分配策略对毛颚类保证足够幼体存活率和维持种群数量具有重要意义。
王崚力 , 胡思敏 , 郭明兰 , 李涛 , 王有军 , 黄晖 , 刘胜 . 三亚湾肥胖软箭虫成体与幼体现场摄食差异研究[J]. 热带海洋学报, 2020 , 39(3) : 57 -65 . DOI: 10.11978/2019092
Chaetognatha is often the most abundant carnivorous zooplankton, and a vital food source for pelagic fish in marine ecosystem. However, their survival strategy of maintaining population structure in tropical coastal waters remains unclear. In this study, molecular method was applied to compare the field diet of juveniles and adults of dominant chaetognath species Flaccisagitta enflata in Sanya Bay. We discuss the nutritional strategies of chaetognaths maintaining substantial population structure from the perspective of resource utilization. The results show that there were 21 and 19 species fed by juveniles and adults, respectively. Copepods, small jellyfish, diatoms, and polychaets were common food groups of both juveniles and adults. Food preferences differed significantly between the juveniles and adults. Small jellyfish made up 59% of the whole diet composition in adults, while copepods became the most abundant food items (60%) in juveniles. The trophic niche of juveniles (5.16) was higher than that of adults (2.89), and trophic overlap between them was low (0.21), suggesting that chaetognaths have obvious dietary shifts during different growth stages. These results reveal that even though adults and juveniles coexist in the same region with large numbers, they can reduce food competition by feeding differentiation. This feeding strategy is of great significance for maxillary species to ensure juvenile survival and maintain substantial population structure.
表1 肥胖软箭虫的克隆序列信息表Tab. 1 Diversity indices of prey organisms in the Flaccisagitta enflata samples analyzed |
| 样品名称 | 克隆数 | OTU数 | Chao1 | 香农多样性指数H’ |
|---|---|---|---|---|
| W3-Jul-a | 55 | 6 | 16 | 0.45 |
| W4-Jul-a | 30 | 7 | 8.5 | 1.55 |
| W9-Jul-a | 38 | 9 | 16.5 | 1.46 |
| 合计 | 123 | 19 | 35.5 | 0.65 |
| W3-Jul-j | 36 | 10 | 13.33 | 1.60 |
| W4-Jul-j | 31 | 8 | 11.33 | 1.15 |
| W9-Jul-j | 47 | 10 | 11 | 1.75 |
| 合计 | 114 | 21 | 32.25 | 2.19 |
注: W3、W4和W9表示站位; Jul表示7月; a表示肥胖软箭虫成体, j表示肥胖软箭虫幼体; 测序样品中成体包含30只肥胖软箭虫, 幼体包含100只肥胖软箭虫。 |
图3 肥胖软箭虫成体与幼体消化道内含物18SrDNA序列组成及其与Genbank中匹配的相似序列所构建的ML(maximum likelihood)进化树 进化树仅显示拓扑结构并以大鼠(Rat sp.)的18S rDNA序列作为外群, 分支处基于1000次重复自举值, 不同宽度的分支代表不同的值, 进化树分支中不同符号的序列标签代表不同站点的样品; 热图值代表样品中食物种类克隆数所占比例 Fig. 3 Composition and Maximum Likelihood (ML) tree of 18S rDNA sequences retrieved from supplied food samples used in Flaccisagitta enflata adults and juveniles diet analysis. Tree is rooted with Rat sp. (M11188.1). Shown at nodes are bootstrap confidence values based on 1000 replicates, and branch thickness depicts the strength of bootstrap support of the corresponding node. The stations of the sample are denoted by different symbols. The thermogram value represents the proportion of food species clones in the sample |
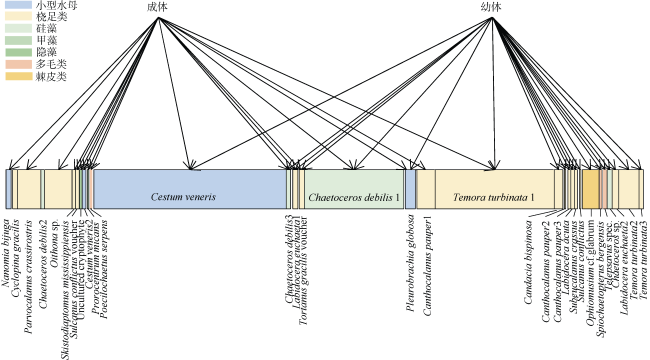
图4 肥胖软箭虫成体与幼体食物来源的二分食物网上层表示不同生长阶段的肥胖软箭虫, 下层代表肥胖软箭虫在OTU水平上的食物来源, 上层与下层的每个连接表示最少一次检测到的摄食事件, 连接的宽度表示检测到的频数, 宽度越宽表示频数越高 Fig. 4 Bipartite networks depicting the main dietary complementary and overlaps of chaetognath adults and juveniles. The upper layer represents Flaccisagitta enflata at different growth stages, and the lower layer represents the food source of F. enfalta at OTU level. Each connection between the upper layer and lower layer represents at least one detected feeding event. The width of the lower layer represents the frequency detected, and wider width represents higher frequency |
| 1 |
蒋建斌, 陆建学 , 2012. 海水桡足类的营养分析及在鱼、虾蟹幼体培育中的应用[J]. 水产养殖, 33(9):1-3.
|
| 2 |
柯志新, 黄良民, 谭烨辉 , 等, 2011. 三亚珊瑚礁分布海区浮游生物的群落结构[J]. 生物多样性, 19(6):696-701.
|
| 3 |
李松, 方金钏 , 1990. 中国海洋浮游桡足类幼体[M]. 北京: 海洋出版社.
|
| 4 |
刘春洋, 王彬, 李轶平 , 等, 2011. 海蜇不同生长阶段的摄食方式和摄食习性[J]. 水产科学, 30(8):491-494.
|
| 5 |
彭瑞冰, 蒋霞敏, 乐可鑫 , 等, 2014. 5种饵料动物的营养成分分析及评价[J]. 水产学报, 38(2):257-264.
|
| 6 |
时翔, 王汉奎, 谭烨辉 , 等, 2007. 三亚湾浮游动物数量分布及群落特征的季节变化[J]. 海洋通报, 26(4):42-49.
|
| 7 |
孙婷婷 , 2018. 基于高通量测序技术的两种水母现场食物研究[D]. 烟台: 烟台大学.
|
| 8 |
王崚力, 郭明兰, 胡思敏 , 等, 2018 -08-10. 一种毛颚类现场食谱检测的分子方法: 中国, CN201810114142.9[P].
|
| 9 |
尹健强, 张谷贤, 黄良民 , 等, 2004a. 三亚湾浮游动物的昼夜垂直移动[J]. 热带海洋学报, 23(5):25-33.
|
| 10 |
尹健强, 张谷贤, 谭烨辉 , 等, 2004b. 三亚湾浮游动物的种类组成与数量分布[J]. 热带海洋学报, 23(5):1-9.
|
| 11 |
郑慧苑, 丘书院 , 1990. 厦门港球型侧腕水母(Pleurobrachia globosa Moser)的生殖[J]. 动物学报, 36(4):393-397.
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}