
珠江口羽流锋浮游植物群落对大气沉降的生态响应
|
周卫文(1992—), 男, 湖南省怀化市人, 博士研究生, 从事海洋生物地球化学研究。E-mail: |
Copy editor: 姚衍桃
收稿日期: 2019-10-15
要求修回日期: 2020-01-06
网络出版日期: 2020-07-27
基金资助
国家自然科学基金项目(41676108)
热带海洋环境国家重点实验室自主研究项目(LTOZZ1705)
热带海洋环境国家重点实验室自主研究项目(LTOZZ1803)
版权
Response of phytoplankton community to atmospheric deposition along Pearl River plume front
Received date: 2019-10-15
Request revised date: 2020-01-06
Online published: 2020-07-27
Supported by
Foundation item: National Natural Science Foundation of China(41676108)
Project of State Key Laboratory of Tropical Oceanography(LTOZZ1705)
Project of State Key Laboratory of Tropical Oceanography(LTOZZ1803)
Copyright
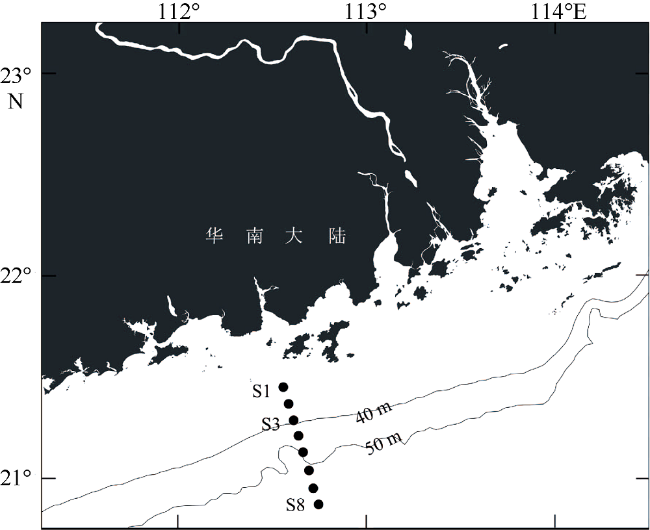
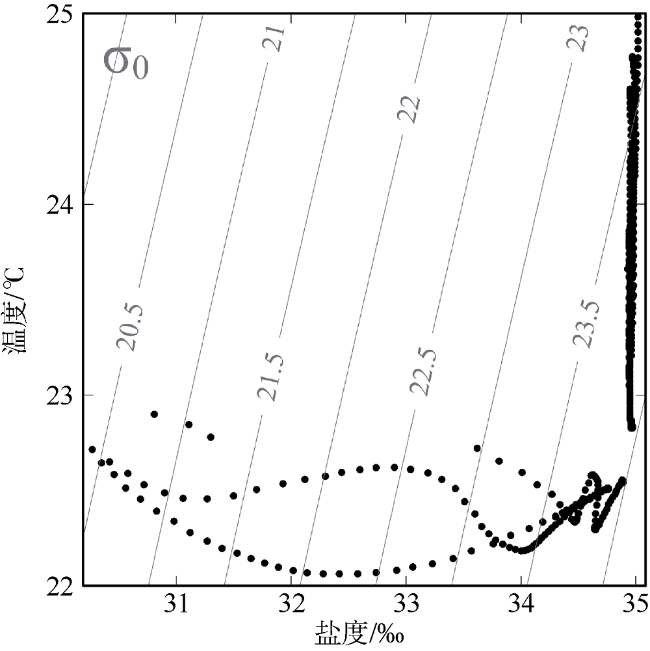
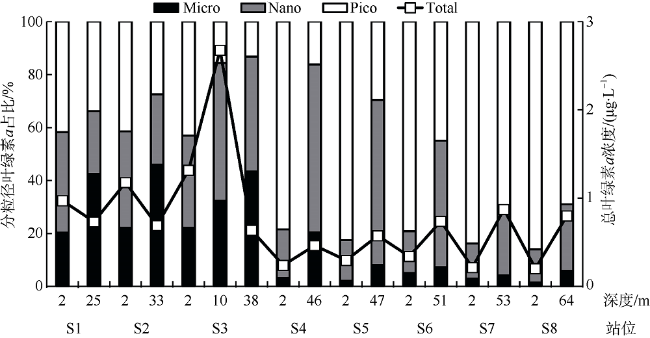
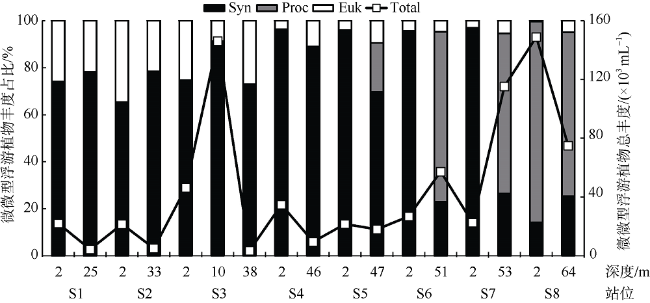
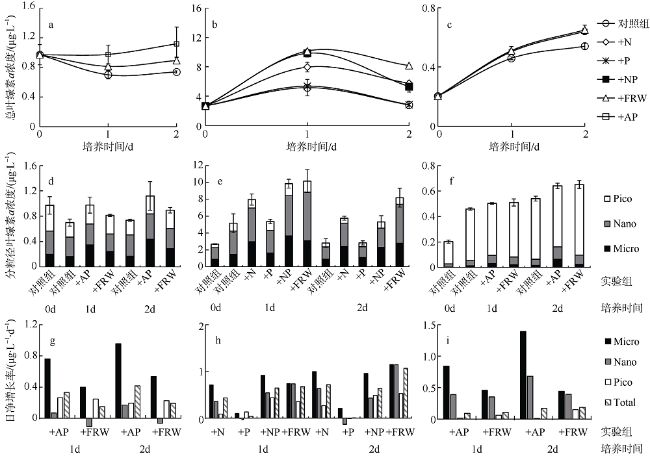
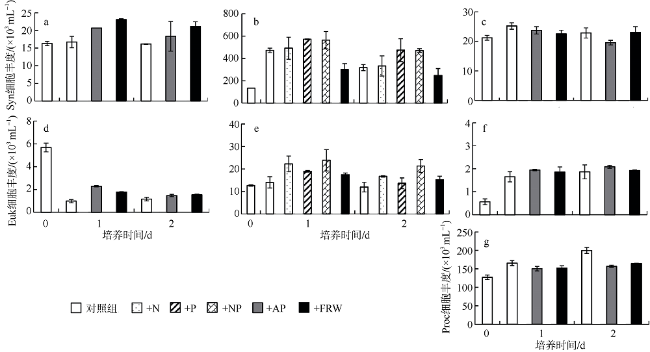
受季风强迫, 珠江淡水羽在南海北部广泛形成羽流锋面, 而浮游植物群落通常在羽流锋面具有较高的生物量。南海北部毗邻珠江三角洲, 陆源输入的大气沉降对该海区生态系统影响显著。利用2019年3月珠江口西部的现场调查数据, 结合羽流锋面浮游植物群落分布的空间差异与大气沉降影响下的浮游植物群落生态学, 分析两者对南海北部陆架区浮游植物粒径群落以及微微型浮游植物的耦合影响。研究表明, 羽流锋面滨侧以小型(Micro)浮游植物群落为主, 锋面海侧以微微型(Pico)浮游植物群落为主, 而锋面区浮游植物群落粒径分布较为均匀, 且生物量高。锋面区域环境因子差异导致浮游植物群落分布呈现较大的空间差异。在锋面区域, 浮游植物群落生长总体受氮限制, 而聚球藻的生长则受磷限制。锋面区域浮游植物群落内部种群对营养盐的需求和响应有所差异。大气干、湿沉降的添加均能够促进不同粒径浮游植物群落的生长: 在锋面滨侧和锋面海侧, 小型浮游植物群落在添加气溶胶颗粒或雨水后比微型(Nano)和微微型浮游植物群落表现出更大的竞争优势, 生长率最高; 而锋面区域浮游植物群落表现出更强的适应性, 小型、微型、微微型浮游植物群落的生长速率均增加且无显著差异。大气沉降颗粒的添加显著促进了锋面系统微微型真核浮游植物与锋面滨侧聚球藻的生长, 在锋面区域以及锋面海侧则抑制了聚球藻和原绿球藻的生长。
周卫文 , 李芊 , 葛在名 , 刘子嘉 , 帅义萍 , 马梦真 . 珠江口羽流锋浮游植物群落对大气沉降的生态响应[J]. 热带海洋学报, 2020 , 39(4) : 50 -60 . DOI: 10.11978/2019101
The Pearl River plume water can form an active plume front along the coastal northern South China Sea. A higher biomass of phytoplankton could exist along the plume front affected by hydrological factors. Atmospheric deposition from land-source input has a significant impact on the ecosystem of the Pearl River Delta zone. A field survey at the western part of the Pearl River Estuary was conducted in March 2019. Substantial variations of size-fractionation phytoplankton community and picoplankton were determined along a section, which affected by the plume front. Meanwhile, atmospheric deposition amendment experiments of three stations, covering different regions of the plume front, were conducted. Results showed that the size-fractionation phytoplankton community was dominated by microphytoplankton at the shore side of the front, picophytoplankton at the sea side of the front, and evenly distributed at the frontal zone with relatively higher biomass. At the frontal zone, the total phytoplankton community was limited by Nitrogen, while the Synechococcus showed a Phosphorus-limitation. Species among the frontal zone could show different nutrients demands. Atmospheric dry or wet deposition could stimulate the growth of three size-fractionation phytoplankton community. The microphytoplankton showed higher growth rate with atmospheric dry or wet amendments at shore side or sea side of the front, while the growth of three size-fractionation phytoplankton community could be stimulated indistinguishably. The addition of atmospheric deposition significantly stimulated the growth of Picoeukaryotes in the frontal system and of Synechococcus at the shore side of the front. Inhibited effects on Synechococcus and Prochlorococcus were found at the frontal zone and sea side of the front.
表1 大气沉降样品的营养盐以及痕量金属浓度Tab. 1 Nutrients and trace metal concentrations of filtered rainwater (FRW) and the leachate solution from aerosol particle (AP) |
| 样品 | 营养盐/(μmol·L-1) | 痕量金属/(nmol·L-1) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DIN | DIP | Al | Cu | As | Cd | Fe | Ni | Co | Cr | Pb | Mn | ||
| 气溶胶滤液(AP) | 6.7 | 0.13 | 104 | 3.6 | 0.76 | 0.38 | 5.0 | 1.4 | 0.14 | 0.25 | 2.3 | 12.2 | |
| 过滤雨水(FRW) | 8.3 | 0.12 | 105 | 3.9 | 2.20 | 0.03 | 3.0 | 1.3 | 0.05 | 0.31 | 1.1 | 3.4 | |
表2 现场营养盐及大气沉降添加的终浓度Tab. 2 Description of different treatments during incubations |
| 实验组 | 添加成分的终浓度 |
|---|---|
| 对照组 | 无添加 |
| +N | 4.2μmol·L-1 |
| +P | 0.25μmol·L-1 |
| +NP | 4.2μmol·L-1(NaNO3)、0.25μmol·L-1(NaH2PO4) |
| +AP | 4.2mg·L-1 |
| +FRW | 10%(体积比例) |
注: +N为加氮组, +P为加磷组, +NP为加氮磷组, +AP为加气溶胶组, +FRW为加过滤雨水组 |
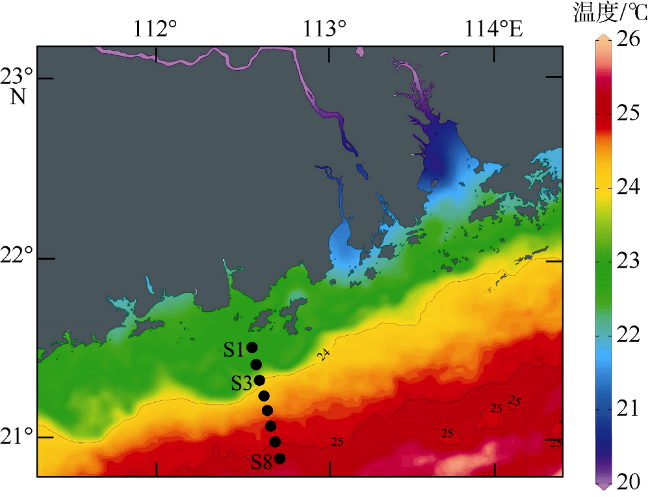
图3 研究区表层水体的温度分布Fig. 3 Distribution of surface temperature in the northern South China Sea |
表3 培养站位不同水深的水文及生物参数Tab. 3 Hydrographic and biological parameters of the incubation stations |
| 参数 | S1 | S3 | S8 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2m | 25m | 2m | 10m | 38m | 2m | 64m | ||||
| 温度/oC | 22.6 | 22.4 | 22.9 | 22.4 | 22.6 | 25.1 | 23.1 | |||
| 盐度/‰ | 30.3 | 34.1 | 32.8 | 33.5 | 34.5 | 34.4 | 34.5 | |||
| 叶绿素a浓度/(µg·L-1) | 1.0 | 0.8 | 1.3 | 2.7 | 0.6 | 0.2 | 0.8 | |||
| 分粒径浮游植物 百分比/% | 小型(Micro) | 20 | 38 | 22 | 30 | 43 | 1 | 6 | ||
| 微型(Nano) | 38 | 32 | 35 | 53 | 44 | 13 | 25 | |||
| 微微型(Pico) | 42 | 30 | 43 | 17 | 13 | 86 | 69 | |||
| 微微型浮游植物细胞丰度/(×103mL-1) | 聚球藻Synechococcus | 16.3 | 3.4 | 119.7 | 133.5 | 31.4 | 21.2 | 19.0 | ||
| 原绿球藻Prochlorococcus | — | — | — | — | — | 127.2 | 52.2 | |||
| 真核浮游植物Picoeukaryotes | 5.7 | 0.9 | 8.4 | 12.6 | 8.0 | 0.6 | 3.7 | |||
注: “—”表示没有检测到该类型浮游植物 |
| [1] |
冯士筰, 李凤岐, 李少菁, 1999. 海洋科学导论[M]. 北京: 高等教育出版社: 1-524(in Chinese).
|
| [2] |
李洁, 经志友, 张偲, 2018. 季风环流影响下的南海海洋细菌多样性特征初探[J]. 热带海洋学报, 37(6):1-15.
|
| [3] |
梁文钊, 唐丹玲, 2017. 南海西部夏季表层浮游植物粒径结构分布特征分析[J]. 热带海洋学报, 36(4):93-101.
|
| [4] |
刘华健, 黄良民, 谭烨辉, 等, 2017. 珠江口浮游植物叶绿素a和初级生产力的季节变化及其影响因素[J]. 热带海洋学报, 36(1):81-91.
|
| [5] |
牟英春, 褚强, 张潮, 等, 2018. 南海浮游植物对沙尘和灰霾添加的响应[J]. 中国环境科学, 38(9):3512-3523.
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}