
塞舌尔角毛藻的形态学再描述和分子系统学分析
|
牛彪彪(1996—), 男, 山西晋城人, 硕士研究生, 从事藻类分类学研究。email: |
Copy editor: 姚衍桃
收稿日期: 2020-06-19
修回日期: 2020-08-08
网络出版日期: 2020-08-18
基金资助
科技基础资源调查专项(2018FY100200)
版权
Morphological and Phylogenetic Studies on Chaetoceros seychellarus
Copy editor: YAO Yantao
Received date: 2020-06-19
Revised date: 2020-08-08
Online published: 2020-08-18
Supported by
Science & Technology Basic Resources Investigation Program of China(2018FY100200)
Copyright
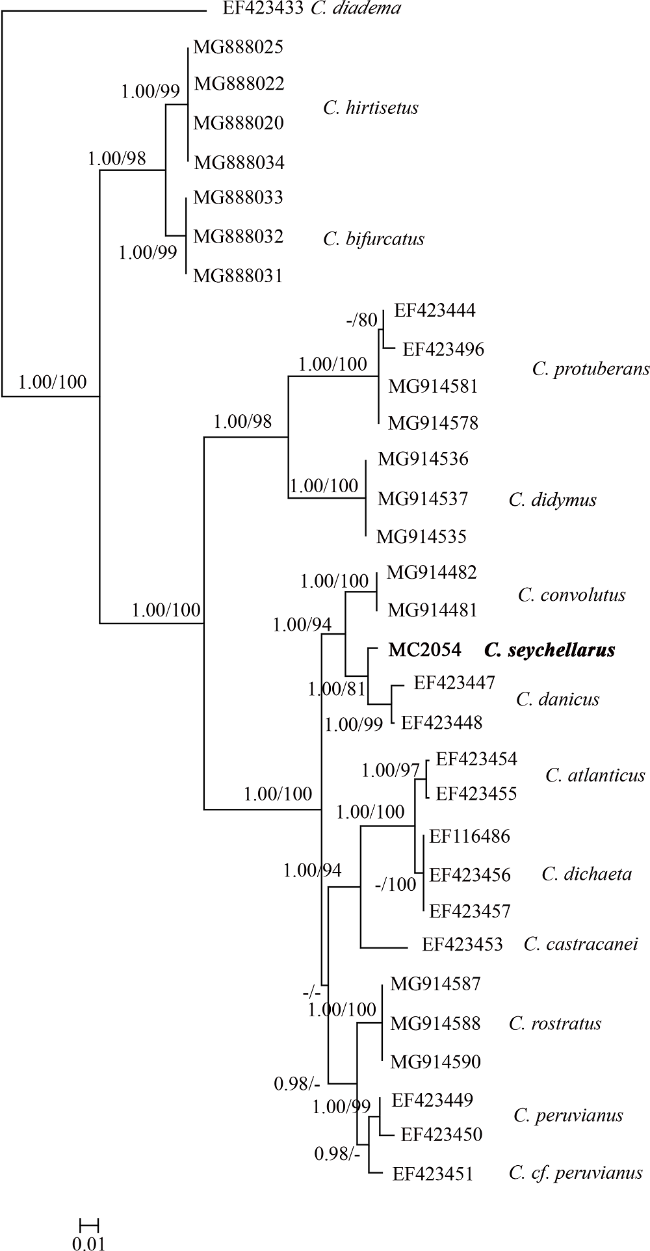
塞舌尔角毛藻(Chaetoceros seychellarus)属于硅藻门角毛藻属有色角毛藻亚属, 为全球罕见种。以往有关塞舌尔角毛藻的研究报道多限于光学显微镜观察, 对其形态学认知仍不完整, 系统学研究也尚属空白。本文从我国台湾海峡和海南岛三亚海域采集了塞舌尔角毛藻, 并在实验室建立了单克隆培养株系, 利用光镜、扫描电镜对其形态学特征进行了较为详尽的观察, 基于核糖体大亚基(large subunit ribosomal DNA)D1~D3区序列, 分析了其系统学信息。塞舌尔角毛藻的典型特征是链状群体直, 窗孔呈矩形, 壳面具有两个唇形突。分子系统树显示, 塞舌尔角毛藻与丹麦角毛藻构成姐妹支, 依据形态学建立的北方组呈现多系起源, 后续有待加以修订。
牛彪彪 , 翟梦怡 , 李扬 . 塞舌尔角毛藻的形态学再描述和分子系统学分析[J]. 热带海洋学报, 2021 , 40(4) : 44 -49 . DOI: 10.11978/2020066
Chaetoceros seychellarus, which belongs to subgenus Phaeoceros of genus Chaetoceros in Bacillariophyta, is a rare species in the world. Present knowledge on the morphology is incomplete, because studies are mainly conducted under LM (light microscope) and lack ultrastructural characteristics under EM (electron microscope). Its systematic information is still unknown. In this study, monoclonal strains of C. seychellarus were established from coastal waters in the southeast China. The morphology was observed using LM and SEM (scanning electron microscope). D1-D3 regions of the large subunit ribosomal DNA (LSU) were amplified to construct the phylogenetic relationship. The key features were straight chains, rectangular apertures and two rimoportulae on each valve that distributed symmetrically along the transapical axis. The phylogenetic tree showed C. seychellarus was a sister to C. danicus. On LSU tree, the section Borealia established based on morphology solely was paraphyletic, which needs revision in further.
Key words: Chaetoceros seychellarus; morphology; phylogeny; section Borealia
表1 塞舌尔角毛藻培养藻株信息Tab. 1 Information of monoclonal strains used in this study |
| 株系编号 | 采样海域 | 采样时间 |
|---|---|---|
| MC2034 | 台湾海峡(117°48'36"E, 22°42'38"N) | 2016年4月2日 |
| MC2054 | 台湾海峡(117°48'36"E, 22°42'38"N) | 2016年4月2日 |
| MC2066 | 台湾海峡(120°20'24"E, 25°33'00"N) | 2016年4月8日 |
| MC2083 | 三亚海域(111°54'00"E, 19°45'94"N) | 2017年4月7日 |
| MC2090 | 三亚海域(111°54'00"E, 19°45'94"N) | 2017年4月7日 |
图1 塞舌尔角毛藻(株系编号: MC2054)a. 群体环面观; b. 群体壳面观; c. 单细胞; d. 上壳面; e. 下壳面; f. 链中细胞(箭头指示壳套与环带连接处); g. 链中相邻壳面(箭头指示唇形突); h. 内壳面观; i. 上壳面; j. 下壳面; k、l. 角毛; m. 角毛截面 Fig. 1 Chaetoceros seychellarus (MC2054). a: Chain in girdle view; b: chain in valve view; c: solitary cell; d: terminal anterior valve; e: terminal posterior valve; f: intercalary cells and arrows showing connection between mantle and bands; g: sibling valves and arrows showing two rimoportulae; h: valve; i: terminal anterior valve; j: terminal posterior valve; k-l: setae structure; and m: setae section |
| [1] |
郭玉洁, 钱树本, 2003. 中国海藻志: 第五卷硅藻门第一册中心纲[M]. 北京: 科学出版社: 379-380(in Chinese).
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}