
海洋动物营养生态位研究方法及其应用
|
李云凯(1981―), 男, 吉林省长春市人, 博士, 从事海洋生态学、稳定同位素生态学、濒危物种保护生物学研究。email: |
Copy editor: 姚衍桃
收稿日期: 2020-07-10
修回日期: 2020-08-29
网络出版日期: 2020-09-02
基金资助
国家自然科学基金项目(31872573)
上海市自然科学基金项目(17ZR1413000)
青岛海洋科学与技术国家实验室海洋渔业科学与食物产出过程功能实验室开放课题(2017-1A03)
农业农村部远洋与极地渔业创新重点实验室开放课题(2019-3)
版权
A review on the methods used in trophic niche studies of marine animals and their applications
Copy editor: YAO Yantao
Received date: 2020-07-10
Revised date: 2020-08-29
Online published: 2020-09-02
Supported by
National Natural Science Foundation of China(31872573)
Natural Science Foundation of Shanghai(17ZR1413000)
Laboratory for Marine Fisheries Science and Food Production Processes, Qingdao National Laboratory for Marine Science and Technology(2017-1A03)
Key Laboratory of Oceanic and Polar Fisheries, Ministry of Agriculture and Rural Affairs(2019-3)
Copyright
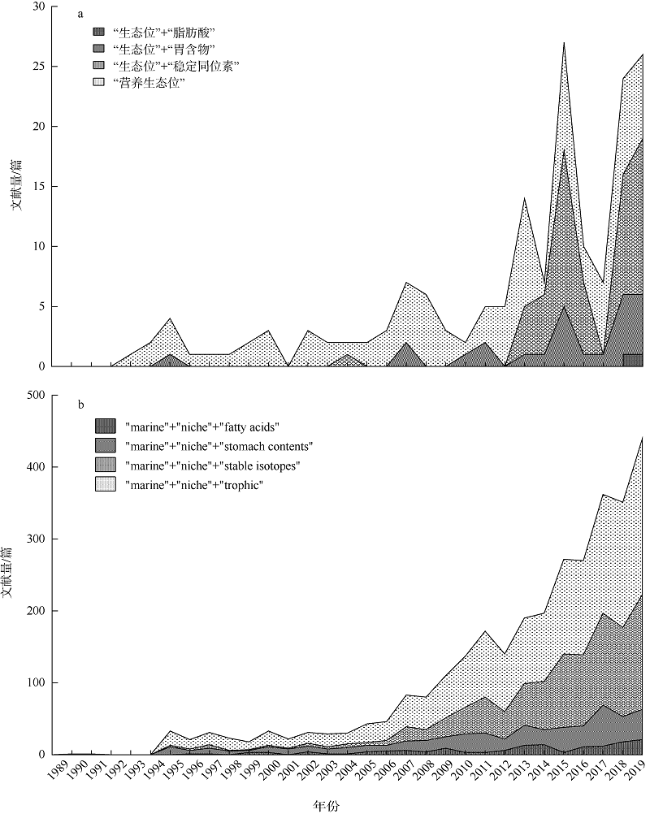
食物资源对海洋动物的生存和繁衍至关重要。海洋动物的营养生态位描述了其在海洋生态系统中的生态地位及功能, 对分析种间关系和资源分配模式、了解群落结构与功能具有重要的指示作用。本文在整理近30年来国内外相关研究的基础上, 系统归纳了当前海洋动物营养生态位研究的主要方法(胃含物分析、整体组织稳定同位素分析、脂肪酸组成分析和特定化合物稳定同位素分析)及其应用领域, 着重归纳总结了量化营养生态位宽度和重叠的生态模型, 并分析了各营养生态位评估方法的优势和局限性。同时在此基础上, 对今后研究方向提出了展望, 以期为我国海洋动物摄食生态学相关研究工作提供参考。
李云凯 , 陈子昂 , 贡艺 , 陈新军 . 海洋动物营养生态位研究方法及其应用[J]. 热带海洋学报, 2021 , 40(4) : 143 -156 . DOI: 10.11978/2020071
Food resources are crucial for the survival and reproduction of marine animals. The trophic niche of marine organisms refers to the ecological roles and functions in marine ecosystem. It plays an important role in studying interspecific relationship, resource partitioning, community structure and function. In this study, by collecting the relevant research on trophic niche of marine animals, we review current research methods (stomach content analysis, bulk tissue stable isotope analysis, fatty acid profiles and compound-specific isotopic analysis) and their applications by focusing on their corresponding ecological models on quantitively evaluating the breadth and overlap of the trophic niche. Moreover, potential development and directions for further studies are presented, with the aim of providing supports for future research on feeding ecology of marine animals.
表1 描述食物组成的单一指数和综合性指数Tab. 1 Single index and comprehensive index in describing food composition |
| 指数类型 | 指数 | 公式 |
|---|---|---|
| 单一指数 | 出现频率(F%) | $F=\frac{某种饵料生物出现次数}{非空胃样品数} \times 100$. |
| 个数百分比(N%) | $N=\frac{某种饵料生物个数}{饵料生物总个数} \times 100$ | |
| 体积百分比(V%) | $V=\frac{某种饵料生物体积}{饵料生物总体积} \times 100$ | |
| 质量百分比(W%) | $W=\frac{某种饵料生物质量}{饵料生物总质量} \times 100$ | |
| 综合性指数 | 相对重要性指数(IRI) | $\text{IRI}=F{\scriptstyle{}^{0}/{}_{0}}(N{\scriptstyle{}^{0}/{}_{0}}+V{\scriptstyle{}^{0}/{}_{0}})$. 或$\text{IRI}=F{\scriptstyle{}^{0}/{}_{0}}(N{\scriptstyle{}^{0}/{}_{0}}+W{\scriptstyle{}^{0}/{}_{0}})$ |
| 绝对重要性指数(AI) | $\text{AI}=F{\scriptstyle{}^{0}/{}_{0}}+N{\scriptstyle{}^{0}/{}_{0}}+W{\scriptstyle{}^{0}/{}_{0}}$ | |
| 优势指数(IP) | ${{I}_{\text{P}}}=\frac{{{V}_{i}}{{F}_{i}}}{\sum{({{V}_{i}}{{F}_{i}})}}$ | |
| 几何重要性指数(GIIj) | $\text{GI}{{\text{I}}_{j}}=\frac{(\sum{{{V}_{i}}{{)}_{j}}}}{\sqrt{n}}$ |
注: Ip中Vi为食物i的体积所占的比例(或W%), Fi为食物i的出现频率; GIIj中Vi为第i个指标(V%、W%、N%), n为所用指标对应的食物总个数 |
表2 基于碳、氮稳定同位素度量营养生态位的定量指标Tab. 2 Quantitative indicators of trophic niche based on carbon and nitrogen stable isotopes |
| 定量指标 | 含义 | 作用 | 参考文献 |
|---|---|---|---|
| δ13C范围 | δ13C最大值与最小值之间的差值 | 可表示基础食物来源的多样性 | Layman et al, 2007; Vaudo et al, 2011; Zhang et al, 2018; 李凡 等, 2018; Costa et al, 2019; 黄佳兴 等, 2019 |
| δ15N范围 | δ15N最大值与最小值之间的差值 | 可表示食物网中的营养长度, 取值范围越大表明营养级越宽泛 | |
| 质心的平均距离 | δ13C、δ15N二维坐标系中, 所有个体到质心的平均距离, 所有个体δ13C、δ15N的均值即为质心坐标 | 代表物种内营养多样性的平均程度 | |
| 最短均值 | δ13C、δ15N二维坐标系中, 相邻两点最短距离的平均值 | 可代表物种在δ13C、δ15N二维坐标系中的整体密度 | |
| 最短均值标准差 | δ13C、δ15N二维坐标系中, 相邻两点间最短距离的标准差 | 可代表物种在δ13C、δ15N二维坐标系中的均匀度, 其值越低表明营养生态位分布更加均匀。相比最短均值, 受样本量大小的影响较小 | |
| 凸包面积 | δ13C、δ15N二维坐标系中, 所有点围成的凸多边形面积 | 可指示食物网中营养多样性的总程度 | |
| 标准椭圆面积 | δ13C、δ15N二维坐标系中, 经协方差矩阵数据处理绘制的椭圆面积 | 量化营养结构和生态位划分, 相比凸包面积, 受样本量影响较小 |
图2 氨基酸代谢过程中的氮同位素分馏[引自Chikaraishi等(2009)]Fig. 2 Nitrogen isotopic fractionation during amino acid metabolism. After Chikaraishi et al (2009) |
表3 胃含物分析、整体组织稳定同位素分析、脂肪酸组成分析及氨基酸单体稳定同位素分析在海洋动物摄食生态学中的比较与应用Tab. 3 Comparison and applications of stomach content analysis, bulk tissue stable isotope analysis, fatty acid profile and compound-specific isotopic analysis of amino acids in trophic ecology study of marine animals |
| 方法 | 优势 | 劣势 | 应用 | 实例 | |
|---|---|---|---|---|---|
| 胃含物分析(SCA) | 操作简单, 能够快速获取真实、具体的摄食信息 | 所需样本量较大, 空胃率较高; 食物消化率直接影响生物食性分析, 且仅能反映生物被捕捞前一段时间内的摄食情况, 存在一定的偶然性 | 食性分析、食物资源时空变化及与生物摄食行为的关系、营养级计算、营养(空间)生态位分析、食物竞争与生物空间分布的关系等 | Hyslop, 1980; Marshall et al, 1997; 韩东燕 等, 2013; Sá-Oliveira et al, 2014 | |
| 稳定同位素技术(SIA) | 可对胃含物食性分析进行补充, 对样本量要求不高, 能够反映生物较长或较短时间内的摄食信息 | 分馏系数和生物基线值易受生物生理、时间、环境变化的影响; 营养级计算不够精确 | 食性分析、食物资源分配及利用、营养级计算、营养生态位分析、物种迁徙洄游及栖息地分析、种间(内)关系分析等 | Post, 2002; Li et al, 2014;Chiu-Werner et al, 2019; Murillo-Cisneros et al, 2019; 盖珊珊 等, 2019; 黄佳兴 等, 2019 | |
| 脂肪酸组成分析(FA) | 能够反映生物一段时间内的摄食信息, 对样本量要求不高, 食性分析较为准确 | 无法计算营养级; 往往使用较多复杂的统计学方法 | 食性分析、食物资源分配及利用、营养生态位分析、种间(内)关系分析等 | Pethybridge et al, 2014; Bierwagen et al, 2019 | |
| 方法 | 优势 | 劣势 | 应用 | 实例 | |
| 氨基酸单体稳定同位素分析(CSIA-AA) | 样本量要求不高, 能够反映生物一段时间内的摄食信息; 营养级计算更为准确 | 前处理较为繁琐; 对氨基酸碳稳定同位素测定较少; 往往作为整体稳定同位素分析的补充手段, 对营养生态位研究尚处于摸索阶段, 没有达成统一标准 | 生物基线确定、营养级计算、营养生态位分析等 | 贡艺 等, 2017; McMahon et al, 2019 | |
| 胃含物分析+稳定同位素技术(SCA+SIA) | 弥补了胃含物“分辨率”受限及仅能反映近期摄食信息的缺点, 食性分析更加准确; 更能揭示食物资源的划分及利用 | 食性分析、食物资源分配及利用、营养级评估、营养生态位分析、种间(内)关系分析等 | Knickle et al, 2014; Garcia et al, 2018; 李凡 等, 2018 | ||
| 稳定同位素技术+脂肪酸分析(SIA+FA) | 弥补了脂肪酸分析无法计算营养级的缺陷; 食性分析结果更准确; 能从不同角度解释营养生态位差异, 从多角度提供更多摄食信息, 研究潜在生态位重叠 | 食性分析、食物资源分配及利用、营养生态位分析、种间(内)关系分析等 | Antonio et al, 2014; Sardenne et al, 2016 | ||
| 稳定同位素技术+氨基酸单体稳定同位素分析(SIA+CSIA-AA) | 解决了生物基线值变化的问题, 计算营养级更为准确; 更能细化营养生态位差异 | 食性分析、食物资源分配及利用、生物基线确定、营养级计算及其时间变化、营养生态位分析、物种迁徙洄游和栖息地分析、种间(内)关系分析等 | Lorrain et al, 2009; Zanden et al, 2013; Ruiz-Cooley et al, 2017; Cherel et al, 2019; Hetherington et al, 2019 | ||
| [1] |
盖珊珊, 赵文溪, 宋静静, 等, 2019. 小黑山岛人工鱼礁区许氏平鲉和大泷六线鱼的营养生态位研究[J]. 生态学报, 39(18):6923-6931.
|
| [2] |
高小迪, 陈新军, 李云凯, 2018. 水生食物网研究方法的发展和应用[J]. 中国水产科学, 25(6):1347-1360.
|
| [3] |
贡艺, 陈玲, 李云凯, 2017. 海洋生态系统稳定同位素基线的选取[J]. 应用生态学报, 28(7):2399-2404.
|
| [4] |
韩东燕, 薛莹, 纪毓鹏, 等, 2013. 胶州湾5种虾虎鱼类的营养和空间生态位[J]. 中国水产科学, 20(1):148-156.
|
| [5] |
黄佳兴, 龚玉艳, 徐姗楠, 等, 2019. 南海中西部海域鸢乌贼中型群和微型群的营养生态位[J]. 应用生态学报, 30(8):2822-2828.
|
| [6] |
黄亮, 吴莹, 张经, 2009. 脂肪酸标志水生生态系统营养关系的研究[J]. 海洋科学, 33(3):93-96.
|
| [7] |
姜亚洲, 林楠, 袁兴伟, 等, 2015. 基于碳、氮稳定同位素技术研究象山港虾虎鱼类营养生态位[J]. 生态学杂志, 34(6):1579-1585.
|
| [8] |
李凡, 徐炳庆, 吕振波, 等, 2018. 莱州湾鱼类群落优势种生态位[J]. 生态学报, 38(14):5195-5205.
|
| [9] |
李㛃, 朱金兆, 朱清科, 2003. 生态位理论及其测度研究进展[J]. 北京林业大学学报, 25(1):100-107.
|
| [10] |
刘屹, 邓竣尹, 谢天资, 等, 2015. 生态位研究进展及干旱河谷生态位研究重点[J]. 四川林业科技, 36(5):16-20.
|
| [11] |
沙永翠, 张培育, 张欢, 等, 2015. 栖息地环境对种群营养生态位的影响——以黄颡鱼为例[J]. 生态学报, 35(5):1321-1328.
|
| [12] |
石焱, 2018. 基于碳氮稳定同位素的闽江口常见鱼类营养生态位季节性变化[D]. 厦门: 集美大学.
|
| [13] |
王凤, 鞠瑞亭, 李跃忠, 等, 2006. 生态位概念及其在昆虫生态学中的应用[J]. 生态学杂志, 25(10):1280-1284.
|
| [14] |
王桂明, 周庆强, 钟文勤, 1996. 内蒙古典型草原4种常见小哺乳动物的营养生态位及相互关系[J]. 生态学报, 16(1):71-76.
|
| [15] |
王娜, 2008. 脂肪酸等生物标志物在海洋食物网研究中的应用——以长江口毗邻海域为例[D]. 上海: 华东师范大学.
|
| [16] |
席晓晴, 鲍宝龙, 章守宇, 2017. DNA条形码在马鞍列岛海域皮氏叫姑鱼胃含物鉴定中的应用[J]. 水产学报, 41(10):1533-1541.
|
| [17] |
薛莹, 金显仕, 2003. 鱼类食性和食物网研究评述[J]. 海洋水产研究, 24(2):76-87.
|
| [18] |
杨璐, 曹文清, 林元烧, 等, 2016. 夏季北部湾九种经济鱼类的食性类型及营养生态位初步研究[J]. 热带海洋学报, 35(2):66-75.
|
| [19] |
杨效文, 马继盛, 1992. 生态位有关术语的定义及计算公式评述[J]. 生态学杂志, 11(2):44-49, 35.
|
| [20] |
原宝东, 闫永峰, 2016. 鹊鸲冬季和春季取食生态位初步研究[J]. 四川动物, 35(3):426-430.
|
| [21] |
钟章成, 1987. 生态位及其在生态学研究中的应用[J]. 资源开发与保护, 3(3):7-12 (in Chinese)
|
| [22] |
周丹, 丛沛桐, 于涛, 等, 1999. 羊草种群生态位的计算方法[J]. 东北林业大学学报, 27(3):48-50.
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}