
北部湾养殖牡蛎体内异养细菌数量及其耐药性研究
|
李炳(1995—), 男, 硕士研究生, 主要从事水产细菌研究. email: |
Copy editor: 姚衍桃
收稿日期: 2020-08-15
修回日期: 2020-10-21
网络出版日期: 2020-10-25
基金资助
国家自然科学基金项目(31902416)
广东省自然科学基金项目(2017A030313112)
广东省现代农业产业技术体系岗位建设专项资金(2020KJ14)
国家贝类产业技术体系建设专项资金(CARS-49)
版权
The number and antibiotic resistance of heterotrophic bacteria in Crassostrea hongkongensis in aquaculture areas of Beibu Gulf
Copy editor: YAO Yantao
Received date: 2020-08-15
Revised date: 2020-10-21
Online published: 2020-10-25
Supported by
National Natural Science Foundation of China(31902416)
Natural Science Foundation of Guangdong Province(2017A030313112)
Funds for Post Construction of Modern Agricultural Industrial Technology System of Guangdong Province(2020KJ14)
Earmarked Fund for Modern Agro-industry Technology Research System(CARS-49)
Copyright
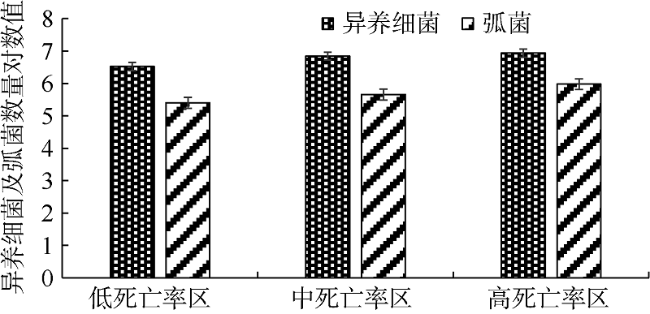
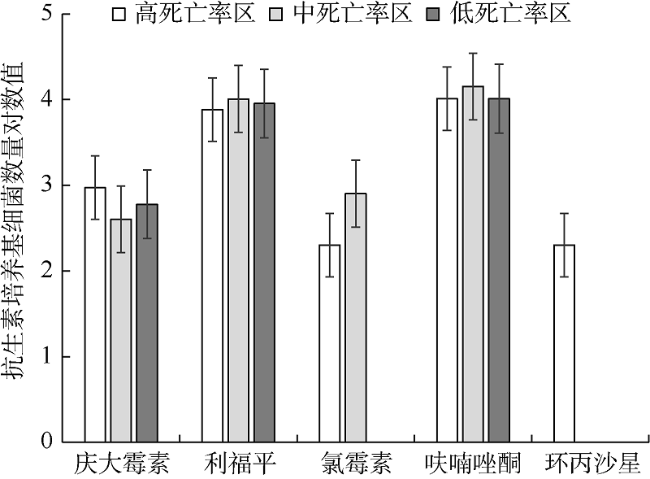
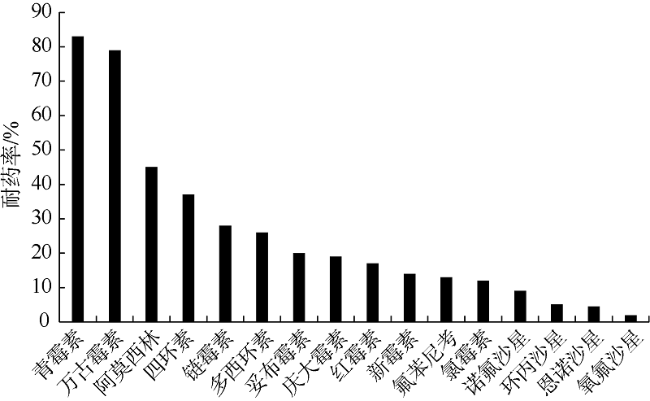
为了探究北部湾养殖区域香港牡蛎(Crassostrea hongkongensis)体内的异养细菌和弧菌数量及其耐药概况变化, 对不同养殖场牡蛎体内的异养细菌进行分离培养, 并统计其数量, 通过药敏纸片扩散等方法研究了细菌的耐药状况。结果显示: 牡蛎在高死亡率养殖环境中体内的异养细菌[(8.6±0.4)×106CFU·g-1]和弧菌[(9.5±0.4)×105CFU·g-1]数量较高, 在中死亡率环境中体内的异养细菌[(6.9±0.2)×106CFU·g-1]和弧菌[(4.5±0.6)×105CFU·g-1]数量次之, 在低死亡率养殖环境中体内的异养细菌[(3.3±0.1)×106CFU·g-1]和弧菌[(2.5±0.6)×105CFU·g-1]数量最低。耐药细菌主要为革兰氏阴性菌, 对β-内酰胺类(青霉素)、糖肽类(万古霉素)的耐药率较高, 对四环素类(四环素、多西环素)的耐药率次之, 对氨基糖苷类(链霉素、庆大霉素、妥布霉素、新霉素)、大环内酯类(红霉素)、喹诺酮类(诺氟沙星、环丙沙星、氧氟沙星、恩诺沙星)的耐药率较低。在高死亡率环境中牡蛎体内的多重耐药菌占79.7%, 其耐药谱型(48种)较广; 在中度死亡率环境中牡蛎体内的多重耐药菌占66.2%, 其耐药谱型为30种; 在低死亡率环境中牡蛎体内的多重耐药菌占58.4%, 其耐药谱型为17种。本文探究了牡蛎死亡率与其体内异养细菌数量和细菌耐药性的关系, 结果显示牡蛎在高死亡率环境中体内的耐药细菌数量多、耐药谱型较广, 低死亡率环境中牡蛎体内的耐药细菌数量较少, 异养细菌数量与牡蛎死亡率呈正相关关系, 两者相关系数为0.996。
李炳 , 王瑞旋 , 张立 , 罗帮 , 牟红莉 , 王江勇 . 北部湾养殖牡蛎体内异养细菌数量及其耐药性研究[J]. 热带海洋学报, 2021 , 40(4) : 70 -83 . DOI: 10.11978/2020092
This study investigated the quantity and antibiotic resistance of heterotrophic bacteria in Crassostrea hongkongensis, which were collected from aquaculture zones with different mortality rates. Heterotrophic bacteria were cultivated and numbered, and their antibiotic resistance was explored by the Kirby-Bauer method. The results indicate that the highest number of heterotrophic bacteria [(8.6±0.4)×10 6CFU·g-1] and Vibrio [(9.5±0.4)×10 5CFU·g-1] occurred in the high-mortality farming zones. The medium number of heterotrophic bacteria [(6.9±0.2)×106CFU·g-1] and Vibrio [(4.5±0.6)×10 5CFU·g-1] occurred in the moderate-mortality farming zones. Heterotrophic bacteria [(3.3±0.1)×106CFU·g-1] and Vibrio [(2.5±0.6)×10 5CFU·g-1] had reached the least abundant in low-mortality farming areas. The antibiotic resistance happened mainly to gram-negative bacteria, especially with the highest resistance rate to β-lactams (penicillin) and glycopeptides (vancomycin), and the second to tetracyclines (tetracycline, doxycycline). The lower resistance rates of antibiotics happened to aminoglycosides (streptomycin, gentamicin, tobramycin, neomycin), macrolides (erythromycin) and quinolones (norfloxacin, ciprofloxacin, ofloxacin, enrofloxacin). Multi-antibiotics-resistant bacteria accounted for 79.7% in high-mortality oysters, with a broad spectrum of antibiotic resistance (48 antibiotics). Multi-antibiotics-resistant bacteria accounted for 66.2% in medium-mortality oysters, with resistance to 30 antibiotics. Heterotrophic bacteria with multi-antibiotics resistance in oysters from the low-mortality area accounted for 58.4%, with resistance to 17 kinds of antibiotic-resistant spectrum. The antibiotic resistance and number of heterotrophic bacteria may be related to the death of oysters. The resistant bacteria of high-mortality oysters were large in number and broad in antibiotic resistance spectrum. Quantity of the antibiotic-resistant bacteria in oysters from the low-mortality area were lower. The results reveal that the number of heterotrophic bacteria is positively correlated with the mortality rate of oysters.
表2 各养殖区牡蛎体内的耐药菌株种属及数量Tab. 2 Classification of antibiotic-resistant strains in oysters from different farming areas |
| 高死亡率区 | 中死亡率区 | 低死亡率区 | |||
|---|---|---|---|---|---|
| 种属 | 数量 | 种属 | 数量 | 种属 | 数量 |
| 弧菌属 (Vibrio) | 14 | 发光杆菌属 (Photobacterium) | 33 | 弧菌属 (Vibrio) | 13 |
| 发光杆菌属 (Photobacterium) | 7 | 弧菌属 (Vibrio) | 25 | 发光杆菌属 (Photobacterium) | 4 |
| 大洋单胞菌属 (Oceanimonas ) | 8 | 海单胞菌属 (Marinomonas) | 5 | 不动杆菌属 (Acinetobacter) | 3 |
| 类香味菌属 (Myroides) | 7 | 大洋单胞菌属 (Oceanimonas) | 2 | 黄杆菌属 (Flavobactrium) | 2 |
| 海单胞菌属 (Marinomonas) | 7 | 希瓦氏菌属 (Shewanella) | 2 | 芽孢杆菌属 (Bacillus) | 1 |
| 藤杆菌属 (Tenacibaculum) | 6 | 芽孢杆菌属 (Bacillus) | 1 | 希瓦氏菌属 (Shewanella) | 1 |
| 希瓦氏菌属 (Shewanella) | 4 | ||||
| 假单胞菌属 (Pseudomonas) | 3 | ||||
| 短波单胞菌属 (Brevundimonas) | 3 | ||||
| 不动杆菌属 (Acinetobacter) | 2 | ||||
| 假黄色单胞菌属 (Pseudoxanthomonas) | 1 | ||||
| 芽孢杆菌属 (Bacillus) | 1 | ||||
| 海盐单胞菌属 (Halomona) | 1 | ||||
表3 低死亡率养殖区牡蛎体内异养细菌的耐药谱Tab. 3 Antibiotic resistance spectrum of heterotrophic bacteria in oysters in low-mortality farming area |
| 序号 | 耐药谱 | 谱型 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PEN | VAN | AMO | TET | STR | DOX | TOB | GEN | ERY | NEO | FLO | CLI | NOR | CIP | ENR | OFL | ||
| 1 | R | S | S | S | S | S | S | S | I | S | S | S | S | S | S | S | H1 |
| 2 | R | I | S | S | S | S | I | S | S | S | S | S | S | S | S | S | H1 |
| 3 | S | S | S | S | R | S | S | S | S | S | S | S | S | S | S | S | H2 |
| 4 | S | S | S | S | I | S | R | S | S | I | S | S | I | I | S | S | H3 |
| 5 | R | R | I | I | S | S | S | S | I | S | S | S | S | S | S | S | H4 |
| 6 | R | R | S | S | S | S | S | S | I | S | S | S | S | S | S | S | H4 |
| 7 | R | R | I | I | I | I | I | S | I | I | S | S | S | S | S | S | H4 |
| 8 | R | S | R | S | S | S | S | S | S | S | S | S | S | S | S | S | H5 |
| 9 | R | S | R | I | I | I | I | S | I | I | S | S | S | S | S | S | H5 |
| 10 | R | S | R | S | S | S | S | S | S | S | S | S | S | S | S | S | H5 |
| 11 | R | R | R | I | S | S | S | S | I | S | S | S | S | S | S | S | H6 |
| 12 | R | R | I | I | S | I | R | S | I | S | S | S | S | S | S | S | H7 |
| 13 | R | R | R | R | S | S | S | S | I | S | S | I | S | S | S | S | H8 |
| 14 | R | R | R | R | S | S | S | S | S | S | S | S | S | S | S | S | H8 |
| 15 | I | S | S | I | R | S | R | R | S | R | S | S | I | S | S | S | H9 |
| 16 | I | S | S | S | R | S | R | R | S | R | S | S | S | S | S | S | H9 |
| 17 | R | R | R | I | I | S | R | S | I | I | I | S | S | S | R | S | H10 |
| 18 | R | R | R | S | R | I | S | S | R | I | S | S | S | S | S | S | H11 |
| 19 | S | S | S | R | S | R | S | S | R | S | R | R | S | S | S | S | H12 |
| 20 | R | R | I | R | I | I | R | R | I | R | S | S | S | S | S | S | H13 |
| 21 | R | R | R | R | R | R | S | S | I | I | S | I | S | S | S | S | H14 |
| 22 | I | S | I | R | S | R | S | S | R | S | R | R | R | S | S | S | H15 |
| 23 | R | I | R | R | R | R | R | I | I | I | I | R | S | I | I | S | H16 |
| 24 | R | R | R | R | R | R | R | R | I | R | R | I | R | R | R | R | H17 |
注: 表中PEN为青霉素、VAN为万古霉素、AMO为阿莫西林、TET为四环素、STR为链霉素、DOX为多西环素、TOB为妥布霉素、GEN为庆大霉素、ERY为红霉素、NEO为新霉素、FLO为氟苯尼考、CLI为氯霉素、NOR为诺氟沙星、CIP为环丙沙星、ENR为恩诺沙星、OFL为氧氟沙星。S、I、R分别表示敏感、中度、耐药3种耐受情况 |
表4 中死亡率养殖区牡蛎体内异养细菌的耐药谱Tab. 4 Antibiotic resistance spectrum of heterotrophic bacteria in oysters in medium-mortality farming area |
| 序号 | 耐药谱 | 谱型 | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PEN | VAN | AMO | TET | STR | DOX | TOB | GEN | ERY | NEO | FLO | CLI | NOR | CIP | ENR | OFL | ||||||
| 1 | I | R | S | S | I | S | I | S | I | S | S | S | S | S | S | S | M1 | ||||
| 2 | R | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | M2 | ||||
| 3 | R | S | S | S | S | S | S | S | I | S | S | S | S | S | S | S | M2 | ||||
| 4 | R | R | S | S | I | S | S | S | I | I | S | S | S | S | S | S | M3 | ||||
| 5 | R | I | S | I | I | I | I | R | I | I | S | S | S | S | S | S | M4 | ||||
| 6 | R | R | S | S | S | S | S | S | S | S | S | S | S | S | S | S | M5 | ||||
| 7 | R | R | S | S | S | S | S | S | I | S | S | S | S | S | S | S | M5 | ||||
| 8 | R | R | I | I | S | I | I | S | I | I | S | S | S | S | S | S | M5 | ||||
| 9 | R | R | I | I | I | I | S | S | I | S | S | S | S | S | S | S | M5 | ||||
| 10 | R | R | S | S | I | I | I | S | I | S | S | S | S | S | S | S | M5 | ||||
| 11 | R | R | S | S | I | S | I | S | I | S | S | S | S | S | S | S | M5 | ||||
| 12 | R | R | S | S | S | I | I | S | I | S | S | S | S | S | S | S | M5 | ||||
| 13 | R | R | S | S | I | S | I | I | I | S | S | S | S | S | S | S | M5 | ||||
| 14 | R | R | S | S | S | S | S | S | I | S | S | S | S | S | S | S | M5 | ||||
| 15 | R | R | S | S | I | S | I | S | I | I | S | S | S | S | S | S | M5 | ||||
| 16 | R | R | S | S | I | S | I | S | I | I | S | S | S | S | S | S | M5 | ||||
| 17 | R | R | S | S | I | S | S | S | I | S | S | S | S | S | S | S | M5 | ||||
| 18 | R | R | S | S | I | I | I | I | I | S | S | S | S | S | S | S | M5 | ||||
| 19 | R | R | S | S | I | I | I | S | I | I | S | S | S | S | S | S | M5 | ||||
| 20 | R | R | S | S | S | I | S | S | I | I | S | S | S | S | S | S | M5 | ||||
| 21 | R | R | S | I | S | S | S | S | I | S | S | S | S | S | S | S | M5 | ||||
| 22 | R | R | S | S | S | I | I | I | I | I | S | S | S | S | S | S | M5 | ||||
| 23 | R | R | S | S | I | S | I | S | I | S | S | S | S | S | S | S | M5 | ||||
| 24 | R | S | R | I | S | S | R | S | S | S | S | S | S | S | S | S | M6 | ||||
| 25 | R | S | R | I | S | S | S | S | I | S | S | S | R | S | I | S | M6 | ||||
| 26 | R | R | R | S | S | S | S | S | I | S | S | S | S | S | S | S | M7 | ||||
| 27 | R | R | R | I | S | I | S | S | I | I | S | S | S | S | S | S | M7 | ||||
| 28 | R | R | R | S | I | I | S | S | I | I | S | S | S | S | S | S | M7 | ||||
| 29 | R | R | R | I | S | I | S | S | I | S | S | S | S | S | S | S | M7 | ||||
| 30 | R | R | R | I | I | I | S | S | I | I | S | S | S | S | S | S | M7 | ||||
| 31 | R | R | R | I | S | I | S | S | I | S | S | S | S | S | S | S | M7 | ||||
| 32 | R | R | R | S | I | S | I | S | I | S | S | S | S | S | S | S | M7 | ||||
| 33 | R | R | R | S | S | S | S | S | I | S | S | S | S | S | S | S | M7 | ||||
| 34 | R | R | R | I | S | I | I | S | I | S | S | S | S | S | S | S | M7 | ||||
| 35 | R | R | R | I | S | I | I | S | I | I | S | S | S | S | S | S | M7 | ||||
| 36 | R | R | R | S | S | S | S | S | I | I | S | S | S | S | S | S | M7 | ||||
| 37 | R | R | I | I | R | S | S | S | I | I | S | S | S | S | S | S | M8 | ||||
| 38 | R | R | S | S | I | S | S | S | R | S | S | S | S | S | S | S | M9 | ||||
| 序号 | 耐药谱 | 谱型 | |||||||||||||||||||
| PEN | VAN | AMO | TET | STR | DOX | TOB | GEN | ERY | NEO | FLO | CLI | NOR | CIP | ENR | OFL | ||||||
| 39 | R | R | S | I | I | I | I | R | I | I | S | S | S | S | S | S | M10 | ||||
| 40 | R | R | S | S | I | S | I | R | I | S | S | S | S | S | S | S | M10 | ||||
| 41 | I | R | S | I | I | I | I | R | I | R | S | S | S | S | S | S | M11 | ||||
| 42 | I | R | S | R | S | R | I | S | I | S | S | S | S | S | S | S | M12 | ||||
| 43 | R | R | S | I | R | S | R | S | I | I | S | S | S | S | S | S | M13 | ||||
| 44 | R | R | S | I | I | R | I | I | I | R | S | S | S | S | I | S | M14 | ||||
| 45 | R | R | I | R | I | R | I | S | I | S | I | I | S | S | I | S | M15 | ||||
| 46 | R | R | S | R | I | R | S | S | I | I | S | S | S | S | S | S | M15 | ||||
| 47 | I | R | S | I | S | R | I | R | I | S | R | S | S | S | I | S | M16 | ||||
| 48 | I | S | S | R | I | R | I | R | R | S | I | S | S | S | I | I | M17 | ||||
| 49 | R | R | R | I | I | I | S | S | R | I | S | S | S | S | S | S | M18 | ||||
| 50 | R | R | R | R | S | I | S | S | I | I | S | S | S | S | S | S | M19 | ||||
| 51 | R | R | R | R | S | I | I | S | I | I | S | S | S | S | S | S | M19 | ||||
| 52 | R | R | R | R | S | S | I | S | I | I | S | S | S | S | S | S | M19 | ||||
| 53 | R | R | R | R | S | S | S | S | I | S | S | S | S | S | S | S | M19 | ||||
| 54 | R | R | R | R | I | I | I | I | I | I | I | S | S | I | I | S | M19 | ||||
| 55 | R | R | S | R | R | R | S | S | S | S | S | S | S | S | S | S | M20 | ||||
| 56 | R | R | S | I | I | R | R | R | I | I | S | S | S | S | S | S | M21 | ||||
| 57 | R | R | R | R | I | R | I | I | I | I | S | S | S | S | I | S | M22 | ||||
| 58 | R | R | R | R | S | R | S | S | I | S | S | S | S | S | S | S | M23 | ||||
| 59 | R | R | R | R | S | R | I | I | I | I | S | S | S | S | S | S | M23 | ||||
| 60 | R | R | R | R | R | R | I | S | I | I | S | S | S | S | S | S | M24 | ||||
| 61 | R | R | R | R | R | R | S | S | S | S | S | S | S | S | S | S | M24 | ||||
| 62 | R | R | R | R | R | R | I | S | I | I | S | S | S | S | S | S | M24 | ||||
| 63 | R | R | R | R | I | R | R | I | I | I | I | I | S | I | I | S | M25 | ||||
| 64 | R | R | S | S | I | R | R | R | I | R | S | S | S | S | S | S | M26 | ||||
| 65 | I | R | I | R | R | R | I | R | I | I | S | S | S | S | R | S | M27 | ||||
| 66 | R | R | R | R | R | I | S | R | R | I | S | I | S | S | S | S | M28 | ||||
| 67 | R | R | R | R | R | R | I | I | I | I | I | S | S | I | R | I | M29 | ||||
| 68 | R | R | I | R | S | R | I | S | R | S | R | S | S | R | I | R | M30 | ||||
注: 表中PEN为青霉素、VAN为万古霉素、AMO为阿莫西林、TET为四环素、STR为链霉素、DOX为多西环素、TOB为妥布霉素、GEN为庆大霉素、ERY为红霉素、NEO为新霉素、FLO为氟苯尼考、CLI为氯霉素、NOR为诺氟沙星、CIP为环丙沙星、ENR为恩诺沙星、OFL为氧氟沙星。S、I、R分别表示敏感、中度、耐药3种耐受情况 |
表5 高死亡率养殖区牡蛎体内异养细菌的耐药谱Tab. 5 Antibiotic resistance spectrum of heterotrophic bacteria in oysters in high-mortality farming area |
| 序号 | 耐药谱 | 谱型 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PEN | VAN | AMO | TET | STR | DOX | TOB | GEN | ERY | NEO | FLO | CLI | NOR | CIP | ENR | OFL | ||||||||
| 1 | R | S | S | S | S | S | S | S | I | S | S | S | S | S | S | S | D1 | ||||||
| 2 | R | S | S | S | S | S | S | S | I | S | S | S | S | S | S | S | D1 | ||||||
| 3 | I | R | S | S | I | S | S | S | I | S | S | S | S | S | S | S | D2 | ||||||
| 4 | I | R | S | S | S | S | S | S | S | S | S | S | S | S | S | S | D2 | ||||||
| 5 | I | R | S | S | I | S | S | S | S | S | S | S | S | S | S | S | D2 | ||||||
| 6 | I | S | S | S | R | S | S | S | I | S | S | S | S | S | S | S | D3 | ||||||
| 7 | S | S | S | S | S | S | S | S | R | S | S | S | S | S | S | S | D4 | ||||||
| 8 | R | R | S | S | I | S | S | I | I | I | S | S | S | S | I | S | D5 | ||||||
| 9 | R | R | S | I | I | S | I | I | I | I | S | S | S | S | S | S | D5 | ||||||
| 10 | R | R | S | S | S | S | S | S | S | S | S | S | S | S | S | S | D5 | ||||||
| 11 | R | R | I | I | I | S | S | S | I | S | S | S | S | S | S | S | D5 | ||||||
| 12 | R | R | S | S | I | S | I | S | I | S | S | S | S | S | S | S | D5 | ||||||
| 13 | R | R | S | S | S | S | S | S | I | S | S | S | S | S | S | S | D5 | ||||||
| 14 | R | R | S | S | I | S | R | I | I | I | S | S | S | S | S | S | D6 | ||||||
| 15 | R | R | S | S | R | S | I | S | S | S | S | S | S | S | S | S | D7 | ||||||
| 16 | R | R | I | R | S | S | S | S | I | S | I | S | S | S | S | S | D8 | ||||||
| 17 | R | R | R | I | S | I | I | I | I | I | S | S | S | S | S | S | D9 | ||||||
| 18 | R | R | S | I | S | S | S | S | R | I | S | S | S | S | S | S | D10 | ||||||
| 19 | R | R | S | I | S | S | R | S | I | S | S | S | S | S | S | S | D11 | ||||||
| 20 | R | R | S | S | I | S | I | I | I | R | S | S | S | S | S | S | D12 | ||||||
| 21 | R | R | S | S | I | S | I | S | R | I | S | S | S | S | S | S | D13 | ||||||
| 22 | I | R | S | R | S | R | S | S | I | S | S | S | S | S | S | S | D14 | ||||||
| 23 | R | R | S | I | I | R | I | S | I | I | S | S | S | S | S | S | D15 | ||||||
| 24 | R | R | S | I | S | R | S | I | R | I | S | S | S | S | S | S | D16 | ||||||
| 25 | R | R | I | R | I | R | S | S | I | S | S | S | I | S | S | S | D17 | ||||||
| 26 | R | S | R | R | S | I | R | S | S | S | S | S | S | S | S | S | D18 | ||||||
| 27 | R | R | R | R | I | I | I | S | I | S | S | S | S | S | S | S | D19 | ||||||
| 28 | R | S | S | S | R | S | S | S | I | S | S | R | R | I | S | S | D20 | ||||||
| 29 | I | S | I | I | R | S | R | R | S | R | S | S | R | I | I | S | D21 | ||||||
| 30 | I | I | I | I | R | S | R | R | S | R | S | S | R | I | I | S | D21 | ||||||
| 31 | R | I | I | I | R | I | R | R | S | R | S | S | I | I | I | S | D22 | ||||||
| 32 | R | R | R | R | I | S | S | S | I | S | I | R | I | S | S | S | D23 | ||||||
| 33 | R | R | R | R | S | S | S | S | I | S | S | S | R | I | S | S | D24 | ||||||
| 34 | R | R | R | R | R | S | I | S | I | S | I | S | S | S | S | S | D25 | ||||||
| 35 | R | R | R | R | R | I | S | I | I | I | I | I | S | S | S | S | D25 | ||||||
| 36 | R | R | R | R | S | R | I | S | I | S | S | S | S | S | S | S | D26 | ||||||
| 37 | I | R | S | I | S | R | R | R | R | S | S | S | S | S | S | S | D26 | ||||||
| 38 | R | I | I | I | R | S | R | R | I | R | S | S | R | I | I | S | D27 | ||||||
| 39 | R | R | R | R | S | S | S | S | R | S | R | R | S | S | S | S | D28 | ||||||
| 40 | R | I | I | I | R | S | R | R | S | R | S | S | R | I | I | S | D29 | ||||||
| 41 | R | S | I | I | R | S | R | R | S | R | S | S | R | I | I | S | D29 | ||||||
| 42 | R | R | R | I | R | S | S | S | R | S | R | S | S | I | S | S | D30 | ||||||
| 43 | R | R | S | R | S | R | S | S | R | S | R | S | S | S | S | S | D31 | ||||||
| 44 | I | R | S | R | R | R | I | S | R | R | S | I | S | S | I | S | D31 | ||||||
| 45 | R | S | I | I | R | I | R | R | I | R | S | S | R | I | I | S | D32 | ||||||
| 序号 | 耐药谱 | 谱型 | |||||||||||||||||||||
| PEN | VAN | AMO | TET | STR | DOX | TOB | GEN | ERY | NEO | FLO | CLI | NOR | CIP | ENR | OFL | ||||||||
| 46 | R | R | R | I | S | I | S | S | R | S | R | R | S | S | S | S | D34 | ||||||
| 47 | I | R | S | R | R | S | R | R | I | S | S | R | S | S | S | S | D35 | ||||||
| 48 | R | S | R | I | R | I | R | R | S | R | S | S | R | I | I | S | D36 | ||||||
| 49 | R | R | R | R | S | I | S | S | R | S | R | R | S | S | I | S | D37 | ||||||
| 50 | R | R | R | R | I | I | S | S | R | S | R | R | S | S | S | S | D37 | ||||||
| 51 | R | R | R | I | R | S | S | S | R | S | R | R | S | S | S | S | D38 | ||||||
| 52 | R | R | R | S | R | S | S | S | R | S | R | R | S | S | S | S | D38 | ||||||
| 53 | R | R | R | R | R | R | I | S | R | I | S | S | S | S | S | S | D39 | ||||||
| 54 | R | R | R | R | R | S | S | S | R | S | R | R | S | S | I | S | D40 | ||||||
| 55 | R | R | S | R | R | I | R | R | R | S | S | R | S | S | S | S | D41 | ||||||
| 56 | I | I | R | R | S | R | I | R | R | S | S | R | R | R | I | I | D41 | ||||||
| 57 | R | I | R | R | R | S | R | R | I | R | S | S | R | R | S | S | D42 | ||||||
| 58 | R | R | R | R | R | R | R | R | R | S | S | I | I | I | S | S | D43 | ||||||
| 59 | R | R | R | I | R | S | R | R | S | R | S | S | R | R | I | S | D44 | ||||||
| 60 | R | R | R | R | R | R | I | S | I | R | S | S | R | R | I | S | D45 | ||||||
| 61 | R | R | R | R | R | R | S | S | R | S | R | R | S | S | S | S | D46 | ||||||
| 62 | R | I | R | R | R | R | R | R | S | R | S | I | R | R | R | I | D47 | ||||||
| 63 | R | R | R | R | R | R | S | S | S | I | R | R | R | R | R | R | D48 | ||||||
| 64 | R | R | R | R | R | R | S | S | S | I | R | R | R | R | R | R | D48 | ||||||
注: 表中PEN为青霉素、VAN为万古霉素、AMO为阿莫西林、TET为四环素、STR为链霉素、DOX为多西环素、TOB为妥布霉素、GEN为庆大霉素、ERY为红霉素、NEO为新霉素、FLO为氟苯尼考、CLI为氯霉素、NOR为诺氟沙星、CIP为环丙沙星、ENR为恩诺沙星、OFL为氧氟沙星。S、I、R分别表示敏感、中度、耐药3种耐受情况 |
| [1] |
陈文博, 王洪军, 李勃, 等, 2012. 养参池底泥弧菌数量变动及微生态制剂对其影响[J]. 水产养殖, 33(10):40-43.
|
| [2] |
董杨, 符一凡, 刘金叶, 等, 2016. 方斑东风螺暴发性疾病的发生与防治[J]. 现代农业科技, (6): 267, 275.
|
| [3] |
李炳, 王瑞旋, 谢燕纯, 等, 2020. 广东沿海香港牡蛎消化道异养菌统计及其耐药性研究[J]. 海洋科学, 44(3):50-58.
|
| [4] |
李兆新, 董晓, 孙晓杰, 等, 2017. 渔业养殖环境中抗生素残留检测与控制技术研究进展[J]. 食品安全质量检测学报, 8(7):2678-2686.
|
| [5] |
李璐璐, 骆延波, 齐静, 等, 2014. CLSI药物敏感性试验标准VET01-A4概述及更新要点[J]. 中国家禽, 36(24):63-66, 69.
|
| [6] |
廉伟, 温海深, 毛玉泽, 等, 2010. 长牡蛎夏季死亡与养殖环境及自身体质关系的初步研究[J]. 渔业科学进展, 31(4):92-100.
|
| [7] |
凌慧, 赵旺, 王瑞旋, 等, 2019. 方斑东风螺养殖环境中细菌数量变动与环境因子的相关性和主成分分析[J]. 生态科学, 38(2):130-137.
|
| [8] |
刘晓靖, 王瑞旋, 凌慧, 等, 2019. 方斑东风螺“急性死亡症”病原及其毒力基因研究[J]. 海洋环境科学, 38(1):7-15.
|
| [9] |
骆向萍, 张静, 刘园园, 2014. 底部微孔增氧技术在海参池塘养殖中应用对比试验[J]. 河北渔业, (4):25-26.
|
| [10] |
毛灿, 苏友禄, 李勇, 等, 2020. 花鲈源杀鱼爱德华菌耐药谱及毒力相关性分析[J]. 中国水产科学, 27(7):846-857.
|
| [11] |
明红霞, 樊景凤, 2015. 气候变化与海洋弧菌疾病的关系[J]. 海洋环境科学, 34(5):792-800.
|
| [12] |
孙永婵, 王瑞旋, 赵曼曼, 等, 2017. 鲍消化道及其养殖水体异养菌的耐药性研究[J]. 南方水产科学, 13(3):58-65.
|
| [13] |
王瑞旋, 耿玉静, 王江勇, 等, 2012. 水产致病菌耐药基因的研究[J]. 海洋环境科学, 31(3):323-328.
|
| [14] |
王瑞旋, 林韵锶, 郭志勋, 等, 2013. 海南陵水企鹅珍珠贝肠道及其养殖水体中异养细菌耐药性研究[J]. 热带海洋学报, 32(6):96-100.
|
| [15] |
魏文娟, 赵姝, 王元, 等, 2020. 副溶血弧菌养殖对虾分离株耐药性及耐药基因分析[J]. 南方水产科学, 16(1):9-16.
|
| [16] |
吴小梅, 林茂, 鄢庆枇, 等, 2015. 美洲鳗鲡及其养殖水体分离耐药菌的多样性和耐药性分析[J]. 水产学报, 39(7):1043-1053.
|
| [17] |
肖慧, 2005. 渤海湾近岸海域的细菌学研究及其在海岸带环境质量评价中的应用[D]. 青岛: 中国海洋大学: 2-4.
|
| [18] |
写腊月, 胡琳琳, 房文红, 等, 2011. 海水养殖源弧菌耐药性调查与分析[J]. 海洋渔业, 33(4):442-446.
|
| [19] |
谢煜, 邢雅丽, 沈钧亮, 等, 2017. 淡水水体中异养细菌丰度研究进展[J]. 浙江万里学院学报, 30(2):89-95.
|
| [20] |
於锋, 罗帮, 陈雪芬, 等, 2016. 广西茅尾海的香港巨牡蛎(Crassostrea hongkongensis)在不同养殖区的性腺发育变化[J]. 渔业科学进展, 37(3):134-142.
|
| [21] |
张锐, 2014. 广东省牡蛎产业发展的公共政策研究[D]. 湛江: 广东海洋大学: 8-11.
|
| [22] |
张颖雪, 苏洁, 叶仕根, 2019. 养殖贝类病原弧菌的耐药性及防治方法研究进展[J]. 江西水产科技, (5):53-56.
|
| [23] |
张占会, 2006. 近江牡蛎致病微生物的筛查与鉴定[D]广州: 暨南大学: 19.
|
| [24] |
钟硕良, 黄万红, 陈月忠, 2012. 厦门海域贝类养殖环境中的大肠菌群和异养细菌[J]. 海洋科学进展, 30(3):357-368.
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
Clinical and Laboratory Standards Institute, 2012. CLSI M100-S22 Performance standards for antimicrobial susceptibility testing; twenty-second informational supplement[S]. Wayne, PA, USA: Clinical and Laboratory Standards Institute.
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}