
海南三亚鹿回头虫黄藻(Effrenium voratum)的形态学和系统发育学研究*
|
龙超(1984—), 男, 湖北省黄冈市人, 博士研究生, 研究方向为海洋生物学。email: |
Copy editor: 姚衍桃
收稿日期: 2020-09-10
修回日期: 2020-11-05
网络出版日期: 2020-11-16
基金资助
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0404)
中国科学院战略性先导科技专项(A类)(XDA13020203)
版权
Morphology and phylogeny of zooxanthellae Effrenium voratum from Luhuitou reef in Sanya, Hainan province
Copy editor: YAO Yantao
Received date: 2020-09-10
Revised date: 2020-11-05
Online published: 2020-11-16
Supported by
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0404)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA13020203)
Copyright
本文对分离于我国海南三亚鹿回头海域的两株虫黄藻SYSC-14-11和SYSC-2-8进行了分类学研究。通过光学显微镜、扫描电子显微镜和分子生物学方法描述了两株藻的形态和系统发育特征, 并与世界其他地理区系的Effrenium属虫黄藻进行了差异性比较, 发现本研究中的两株虫黄藻的形态和系统发育特征与Effrenium属虫黄藻模式种Effrenium voratum基本一致, 推测本文中的两株Effrenium属虫黄藻均为E. voratum。本研究丰富了我国热带海域虫黄藻的物种多样性, 为完善我国的虫黄藻种质资源奠定了基础。
关键词: 虫黄藻; 共生藻科; Effrenium属; 形态学; 系统发育学
龙超 , 罗肇河 , 韦章良 , 杨芳芳 , 李茹 , 龙丽娟 . 海南三亚鹿回头虫黄藻(Effrenium voratum)的形态学和系统发育学研究*[J]. 热带海洋学报, 2021 , 40(4) : 35 -43 . DOI: 10.11978/2020102
In this study, we analyzed taxonomy of two strains (SYSC-14-11 and SYSC-2-8) of zooxanthellae isolated from the Luhuitou reef region in Sanya. Morphological characteristics were determined by light microscope and scanning electron microscope, while phylogenetic analyses were constructed based on the sequences of the LSU rDNA and the internal transcribed spacer sequence (ITS2). The results showed that the morphological and phylogenetic characteristics of the two strains in the present study were basically consistent with Effrenium voratum reported in other geographical regions of the world. This study enriches the species diversity of zooxanthellae in the tropical coral reef ecosystems, and provides a solid foundation for further utilizing the resources of zooxanthellae.
Key words: Zooxanthellae; Symbiodiniaceae; Effrenium; morphology; phylogeny
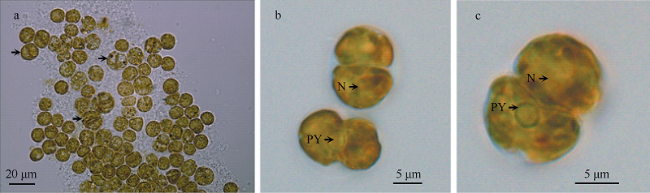
图1 藻株SYSC-14-11的光镜照片a. 不动的球状细胞和运动的葫芦状细胞, 可见二分裂现象; b. 细胞背面观和侧面观, 可见蛋白核及细胞核分布; c. 细胞腹面观, 可见蛋白核及细胞核分布。N: 细胞核; PY: 球状蛋白核 Fig. 1 Light microscope (LM) images of strains SYSC-14-11. a) Immobile globular cells and moving gourd-like cells, including binary dividing cells; b) dorsal view and side view, with pyrenoid and nucleus; and c) ventral view, with pyrenoid and nucleus. N: nucleus; PY: pyrenoid |
图2 藻株SYSC-14-11的扫描电镜照片a. 顶面观, 可见单线长甲板型顶沟复合体(Elongate Apical Vesicle, EAV)和5块顶板, 可见连接板(x); b. 顶面观, 可见4块顶板; c. 顶侧腹面观, 可见横沟与横沟板(C)、纵沟与纵沟板(S); d. 背面观, 可见上锥部、横沟和下锥部; e. 底侧腹面观, 可见横沟和纵沟; f. 底面观, 可见下锥部、纵沟板(S)和沟后板(S.p.) Fig. 2 Scanning electron microscope (SEM) images of strains SYSC-14-11. a) Apical view showing Elongate Apical Vesicle, five apical plates and x plate; b) apical view showing four apical plates; c) apical-ventral view showing cingulum, cingulum plates, sulcus and sulcus plates; d) dorsal view showing episome, cingulum and hyposome; e) antapical-ventral view showing cingulum and sulcus; and f) antapical view showing hyposome, sulcus plates and posterior sulcal plate |
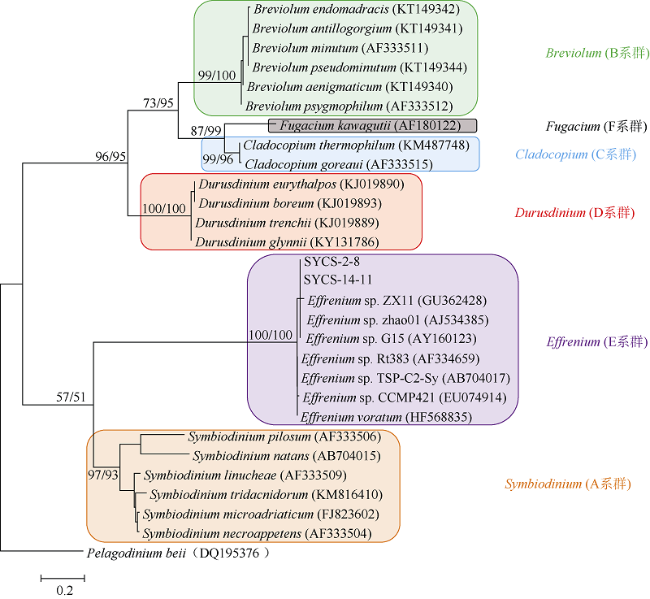
图3 基于核糖体大亚基(LSU)序列构建的系统发育进化树系统发育进化树采用最大似然法(ML)和邻接法(NJ)建立, 左侧node value值为ML, 右侧node value值为NJ。P. beii为外类群, 可信度检测1000次 Fig. 3 Maximum likelihood and neighbor-Joining phylogenetic tree based on LSU sequences with P. beii as outgroup. Bootstrap test (1000 replicates) are shown next to the branches as ML/NJ. For ML, mode/method: Kimura 2-parameter model; Rate among Sites: Gamma distributed with Invariant sites (G + I); No of Discrete Gamma Categories: 5; Gaps/Missing Data Treatment: Partial deletion; Site coverage cutoff: 95%; Select codon positions: 1st + 2nd + 3rd + Noncoding sites; ML Heuristic Method: nearest-neighbor-interchange (NNI). For NJ, mode/method: Kimura 2-parameter model; Substitutions to Include: d Transition + Transversions; Rate among Sites: Gamma Distributed (G); Gamma Parameter: 5, Pattern among Lineages: Same (Homogeneous); Gaps/Missing Data Treatment: Partial deletion; Site coverage cutoff: 95%; Select codon positions: 1st + 2nd + 3rd + Noncoding sites |
图4 基于核糖体转录间隔区(ITS2)序列构建的系统发育进化树系统发育进化树采用最大似然法(ML)和邻接法(NJ)建立, 左侧node value值为ML, 右侧node value值为NJ。P. beii为外类群, 可信度检测1000次 Fig. 4 Maximum likelihood and neighbor-Joining phylogenetic tree based on ITS2 region with P. beii as outgroup. Bootstrap test (1000 replicates) are shown next to the branches as ML/NJ. For ML, mode/method: Kimura 2-parameter model; Rate among Sites: Gamma distributed with Invariant sites (G + I); No of Discrete Gamma Categories: 5; Gaps/Missing Data Treatment: Partial deletion; Site coverage cutoff: 95%; Select codon positions: 1st + 2nd + 3rd + Noncoding sites; ML Heuristic Method: nearest-neighbor-interchange (NNI). For NJ, mode/method: Kimura 2-parameter model; Substitutions to Include: d Transition + Transversions; Rate among Sites: Gamma Distributed (G); Gamma Parameter: 5; Pattern among Lineages: Same (Homogeneous); Gaps/Missing Data Treatment: Partial deletion; Site coverage cutoff: 95%; Select codon positions: 1st + 2nd + 3rd + Noncoding sites |
| [1] |
张跃环, 肖述, 张扬, 等, 2018. 一种砗磲虫黄藻的分离纯化、离体培养及规模化生产方法: 中国, 201710979322.9[P] (in Chinese).
|
| [2] |
赵振鲁, 刘甲星, 张跃环, 等, 2019. 离体培养的虫黄藻(Symbiodinium voratum)对温度和光照的生理响应[J]. 海洋与湖沼, 50(2):316-323.
|
| [3] |
朱霞, 甄毓, 于志刚, 2011. 一株分离自胶州湾的裸甲藻形态相似种的分子系统学研究[J]. 海洋学报, 33(1):153-162.
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}