
全球气候变化对红树林生态系统的影响、挑战与机遇
|
王友绍(1963—), 男, 博士, 山东省临沂市人, 教授, 博士生导师, 主要从事海洋环境生态学研究。email: |
Copy editor: 林强
收稿日期: 2020-11-16
要求修回日期: 2021-01-07
网络出版日期: 2021-01-08
基金资助
国家自然科学基金项目(U1901211)
国家自然科学基金项目(41876126)
国家自然科学基金项目(41430966)
国家自然科学基金项目(4117610)
国家自然科学基金项目(41076070)
国家重点研发计划项目(国家科技基础资源调查专项)(2017FY100700)
中国科学院A类战略性先导科技专项(XDA23050200)
中国科学院A类战略性先导科技专项(XDA13010500)
中国科学院A类战略性先导科技专项(XDA13020503)
国际伙伴计划(133244KYSB20180012)
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0305)
版权
Impacts, challenges and opportunities of global climate change on mangrove ecosystems
Copy editor: LIN Qiang
Received date: 2020-11-16
Request revised date: 2021-01-07
Online published: 2021-01-08
Supported by
National Natural Science Foundation of China(U1901211)
National Natural Science Foundation of China(41876126)
National Natural Science Foundation of China(41430966)
National Natural Science Foundation of China(4117610)
National Natural Science Foundation of China(41076070)
National Key Research and Development Plan (Science & Technology Basic Resources Investigation Program of China)(2017FY100700)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA23050200)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA13010500)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA13020503)
International Partnership Program of Chinese Academy of Sciences(133244KYSB20180012)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0305)
Copyright
红树林是以红树植物为主体的常绿灌木或乔木组成的潮滩湿地木本植物群落, 具有“四高”特性(高生产力、高归还率、高分解率和高抗逆性)的典型海洋生态系统; 目前, 全球约有红树林1700万公顷, 主要分布在南北半球25℃等温线内。红树林生态系统的净初级生产力高达2000gC·m-2·a-1, 具有高强度的物质循环、能量流动以及丰富的生物多样性, 对热带、亚热带海洋生态系统的维持与发展起到关键作用, 并在全球变化过程中扮演着十分重要的角色。近30年来, 全球气候变化已引起了国内外学者的极大关注。红树林生态系统位于热带、亚热带海岸潮间带, 是一个脆弱的、敏感的生态系统, 也是首先受全球气候变化影响的典型海洋生态系统之一。作为全球海岸带地区应对全球气候变化最为重要的生态屏障之一, 气候变化将严重影响着全球红树林的生存和分布方式。本文将从全球变暖、海平面上升、大气中CO2浓度的增加和极端天气4个主要方面, 揭示全球气候变化对红树林生态系统的影响与变化特征, 阐述红树林对全球变暖、海平面上升、大气中CO2浓度增加和极端天气响应与适应的生态学机制, 并简要概述了红树林在减缓全球气候变化危害中的重要作用。全球气候变化也将为红树林的研究、保护和发展带来机遇与挑战。
王友绍 . 全球气候变化对红树林生态系统的影响、挑战与机遇[J]. 热带海洋学报, 2021 , 40(3) : 1 -14 . DOI: 10.11978/YG2020006
Mangroves are salt-tolerant plants of tropical and subtropical intertidal regions distributed mainly between latitudes 25°S and 25°N globally. They have high productivity, high return rate, high decomposition rate, and high resistance, which cover roughly 60%-75% of the world’s tropical coastlines. Mangroves are distributed over more than 118 countries with a total area near 17 million hectares in the world, and their net primary productivity is up to 2000 gC·m-2·a-1. They form a widespread ecosystem and also play an important role in the process of global change. Global climate change has aroused great concerns in the last three decades. Mangroves are a vulnerable and eco-sensitive ecosystem along tropical and subtropical intertidal shores that are regulated by both lands and oceans. They are one of the typical marine ecosystems that can be affected by global climate change. As an important ecological barrier along the coast, global climate change will also affect the survival and distribution of mangroves in the world. The present paper briefly reviews the impacts of global climate change on mangroves in terms of global warming, sea-level rise, atmospheric CO2 concentration increase, and extreme weather. The ongoing global climate change will bring great challenges and opportunities to research, maintenance, and development of mangroves in the future.
图1 浙江温州(霓屿, 左)和苍南(右)北引红树林试验示范区(王友绍, 2019)Fig. 1 Mangroves areas in Wenzhou (Niyu, left) and Cangnan (right) of Zhejiang province, China (Wang, 2019) |
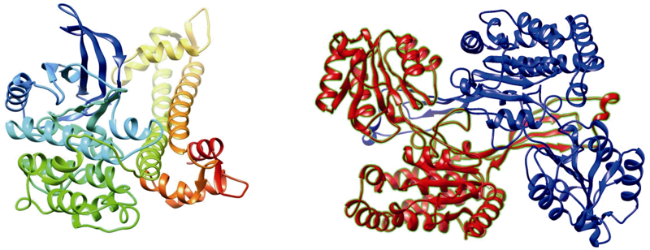
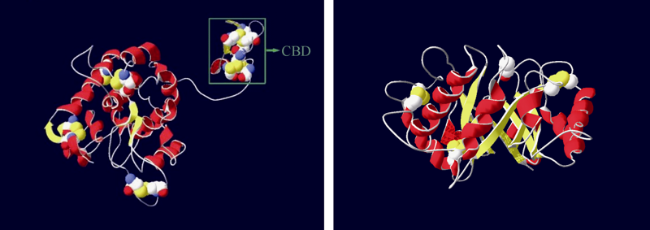
图2 Am CDPK(左)(Liu et al, 2020a)和Am P5CS蛋白(右)(Liu et al, 2020b)三维结构Fig. 2 Three-dimensional structures of Am CDPK protein (left, Liu et al, 2020a) and Am P5CS protein (right, Liu et al, 2020b) |
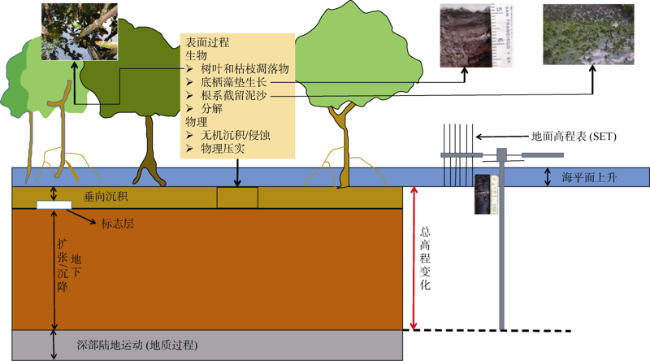
图4 红树林如何适应海平面上升(改自Krauss et al, 2014)Fig. 4 Schematics showing how mangrove forests adjust to rising sea level (modified from Krauss et al, 2014) |
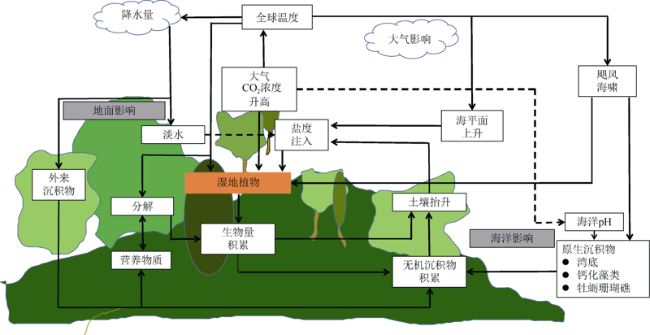
图5 全球气候变化对滨海湿地生态系统影响因子概念模型(改自McKee et al, 2012)Fig. 5 Conceptual model of global change factors influencing coastal wetlands (odified from McKee et al, 2012) |
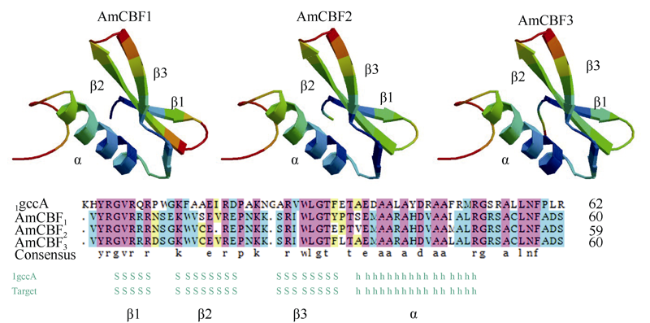
图6 白骨壤CBF/DREB1转录因子AP2结合域的结构与组成(Peng et al, 2013)Fig. 6 Sequence and molecular models of the AP2 domain of CBF/DREB transcription factors in Avicennia marina. After Peng et al (2013) |
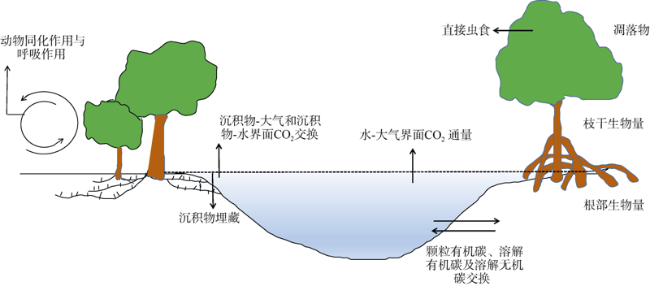
图7 红树林碳收支情况: 初级生产和各种汇(Bouillon et al, 2008; 王友绍, 2019)Fig. 7 Mangrove carbon budgets considered: primary production (litter fall, wood and root production) and various sink terms (Bouillon et al, 2008; Wang, 2019) |
图8 气候变化对红树林影响(改自Faridah-Hanum et al, 2014)Fig. 8 Climate change effects on mangroves (modified from Faridah-Hanum et al, 2014) |
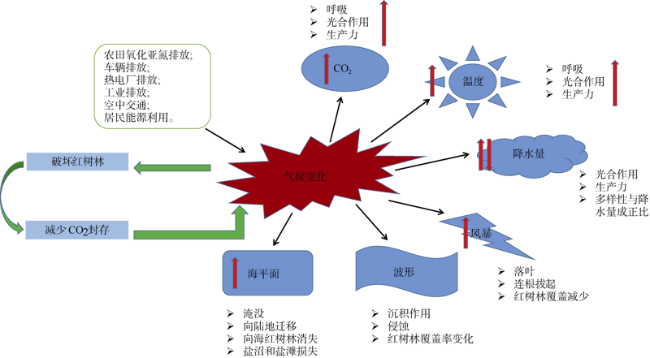
图9 具有多重抗逆性能的桐花树几丁质酶I(左)、III(右)型基因(Wang et al, 2015a, b)Fig. 9 The Aegiceras corniculatum I chitinase gene (Left: AcCHI I) and III chitinase gene (Right: AcCHI III) for anti multi-stress properties (Wang et al, 2015a, b) |
图10 红树林对极端环境变化的响应与适应生态机理 “四高”特性模式图(王友绍, 2019)Fig. 10 Conceptual ecological model of mangroves for responses and adaptation to extreme environments, namely, mangroves having four high characteristics of the ecosystems in the world (Wang, 2019) |
| [1] |
陈鹭真, 王文卿, 张宜辉, 等, 2010. 2008年南方低温对我国红树植物的破坏作用[J]. 植物生态学报, 34(2):186-194.
|
| [2] |
陈玉军, 郑德璋, 廖宝文, 等, 2000. 台风对红树林损害及预防的研究[J]. 林业科学研究, 13(5):524-529.
|
| [3] |
黄晓林, 彭欣, 仇建标, 等, 2009. 浙南红树林现状分析及开发前景[J]. 浙江林学院学报, 26(3):427-433.
|
| [4] |
蒋高明, 韩兴国, 林光辉. 1997. 大气CO2浓度升高对植物的直接影响—国外十余年来模拟实验研究主要手段及基本结论[J]. 植物生态学报, 21(6):489-502.
|
| [5] |
李玫, 廖宝文, 管伟, 等, 2009. 广东省红树林寒害的调查[J]. 防护林科技, (2):29-31.
|
| [6] |
林鹏, 沈瑞池, 卢昌义, 1994. 六种红树植物的抗寒特性研究[J]. 厦门大学学报(自然科学版), 33(2):249-252.
|
| [7] |
林鹏, 1997. 中国红树林生态系[M]. 北京: 科学出版社.
|
| [8] |
彭亚兰, 王友绍, 2014. 红树植物桐花树EF1A基因的克隆与表达分析[J]. 生态科学, 33(4):704-712.
|
| [9] |
王友绍, 2013. 红树林生态系统评价与修复技术[M]. 北京: 科学出版社.
|
| [10] |
王友绍, 2019. 红树林分子生态学[M]. 北京: 科学出版社.
|
| [11] |
杨盛昌, 林鹏, 1998. 潮滩红树植物抗低温适应的生态学研究[J]. 植物生态学报, 22(1):60-67.
|
| [12] |
叶笃正, 1999. 中国的全球变化与可持续发展研究[J]. 地球科学进展, 14(4):317-318.
|
| [13] |
叶勇, 卢昌义, 郑逢中, 等, 2004. 模拟海平面上升对红树植物秋茄的影响[J]. 生态学报, 24(10):2238-2244.
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
FAO, 2007. The world's mangroves 1980-2005[R]. Rome: Food and Agriculture Organization of the United Nations.
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}