
盐度对中国鲎幼鲎生长、蜕壳、Na+-K+-ATP酶活性、免疫指标和抗氧化能力的影响
|
董兰芳(1987—), 女, 硕士, 副研究员, 主要从事水生动物营养学与养殖水环境生态学研究。email: |
Copy editor: 林强
收稿日期: 2021-08-20
修回日期: 2021-10-20
网络出版日期: 2021-10-25
基金资助
广西自然科学基金项目(2018GXNSFBA281071)
广西科学院创新团队启动经费项目(CQ-D-2410)
Effects of salinity on growth, molting, Na+-K+-ATP enzyme activities, immune index, and antioxidantion of juvenile Chinese horseshoe crab, Tachypleus tridentatus
Copy editor: LIN Qiang
Received date: 2021-08-20
Revised date: 2021-10-20
Online published: 2021-10-25
Supported by
Guangxi Natural Science Foundation(2018GXNSFBA281071)
Start-up Fund for Innovation Team of Guangxi Academy of Sciences(CQ-D-2410)
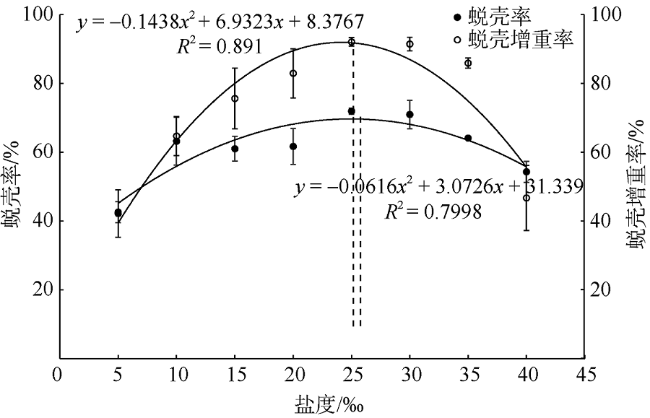
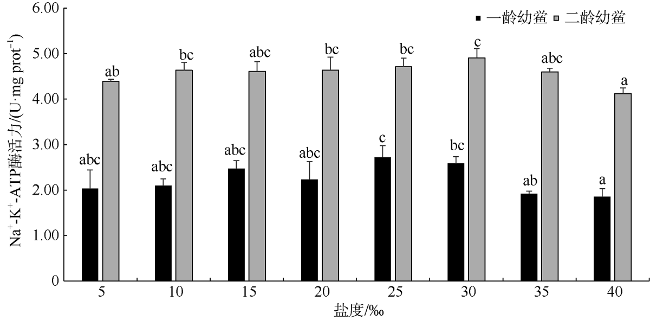
人工增殖放流是恢复鲎资源最有效、最迅速的方法, 而了解和掌握环境因子对中国鲎(Tachypleus tridentatus)幼鲎生长状况的影响规律, 进而选择适宜放流的时间和海区, 是保证人工放流得以成功的关键。本文在实验室条件下, 研究了不同盐度(5‰、10‰、15‰、20‰、25‰、30‰、35‰、40‰)对中国鲎幼鲎成活率、蜕壳率、蜕壳增重率、Na+-K+-ATP酶活性、免疫酶活性、抗氧化酶活性等的影响, 探讨了不同盐度水平下中国鲎幼鲎生长、蜕壳、渗透调节能力、机体免疫力和抗氧化能力等的变化。养殖试验持续56d, 结果表明: 不同盐度对中国鲎幼鲎的成活率、蜕壳率、二龄幼鲎均重及蜕壳增重率均有显著影响(P<0.05), 且均随盐度升高呈先升高后降低的趋势; 蜕壳率和蜕壳增重率与盐度的回归分析均表明, 中国鲎幼鲎蜕壳与生长的最适盐度分别为24.10‰和24.94‰; 一龄和二龄幼鲎的Na+-K+-ATP酶活性均随盐度的升高呈先显著升高后显著降低趋势(P<0.05); 35‰和40‰盐度试验组一龄幼鲎的酸性磷酸酶(acid phosphatase, ACP)活性显著高于其他试验组, 而5‰盐度试验组一龄幼鲎的碱性磷酸酶(alkaline phosphatase, AKP)活性显著低于25‰试验组(P<0.05) ; 盐度对二龄幼鲎的ACP、AKP和溶菌酶(lysozyme, LZM)活性均没有显著影响(P>0.05); 35‰和40‰试验组一龄幼鲎的过氧化氢酶(catalase, CAT)活性显著高于盐度较低试验组(P<0.05); 二龄幼鲎的谷胱甘肽过氧化物酶(glutathione peroxidase)活性随着盐度的升高而升高, 盐度10‰试验组显著低于30‰和40‰试验组(P<0.05)。研究结果显示盐度对中国鲎幼鲎生长、蜕壳、Na+-K+-ATP酶活性、免疫指标和抗氧化能力均有显著影响, 蜕壳最适宜的盐度在24‰~25‰左右, 盐度过高或过低都将引起幼鲎生长率和成活率降低, 渗透调节能力、免疫力和抗氧化力显著下降。
董兰芳 , 许明珠 , 刘海娟 , 曾梦清 , 陈瑞芳 , 李世才 . 盐度对中国鲎幼鲎生长、蜕壳、Na+-K+-ATP酶活性、免疫指标和抗氧化能力的影响[J]. 热带海洋学报, 2022 , 41(3) : 156 -163 . DOI: 10.11978/2021106
Artificial proliferation and release is the most effective and rapid method to restore Limulus resources. Understanding and grasping the influence of environmental factors on the growth of juvenile Chinese horseshoe crab (Tachypleus tridentatus), and then choosing a suitable time and sea area for release are the key steps to ensure the success of manual release. Under laboratory conditions, we studied the effects of different salinity (5‰, 10‰, 15‰, 20‰, 25‰, 30‰, 35‰, and 40‰) on the survival rate, molting rate, molting weight gain rate, Na+-K+-ATPase activity, immune enzyme activity, and antioxidant enzyme activity of juvenile Chinese horseshoe crab. The changes of growth, molting, osmotic adjustment, immunity, and antioxidation of the juvenile horseshoe crab at different salinity levels were recorded. The culture experiment lasted for 56 days. The results showed that different salinity levels significantly affected the survival rate, molting rate, average weight of the 2nd instar horseshoe crabs, and molting weight gain rate of the juvenile horseshoe crab (P< 0.05), which increased at first and then decreased with increasing salinity. Regression analysis of molting rate, molting weight gain rate and salinity showed that the optimum salinity for molting and growth of young horseshoe crab were 24.10‰ and 24.94‰, respectively. The Na+-K+-ATPase activities of the 1st and 2nd instar horseshoe crabs increased at first and then decreased significantly with the increase of salinity (P< 0.05). The acid phosphatase (ACP) activities of the 1st instar larvae in the 35‰ and 40‰ test groups were significantly higher than those in the other test groups, while the alkaline phosphatase (AKP) activities of the 5‰ test group were significantly lower than those in the 25‰ test group (P< 0.05). The activities of ACP, AKP and lysozyme (LZM) of the 2nd instar larvae did not change significantly under different salinity conditions (P> 0.05). The catalase (CAT) activities of the 1st instar larvae in 35‰ and 40‰ test groups were significantly higher than those of low-salinity test groups (P< 0.05). And the glutathione peroxidase (GSH-Px) activities of the 2nd WHAT tended to increase with the increase of salinity, and those in 10‰ test group were significantly lower than those in 35‰ and 40‰ test groups (P< 0.05). We show that salinity had a significant effect on growth, molting, Na+-K+-ATPase activity, immune index, and antioxidation of the juvenile horseshoe crab. The optimum salinity for molting was between 24‰ and 25‰. Too high or too low salinity will cause the growth rate, survival rate of juvenile silver carp, the osmotic adjustment ability, immunity, and antioxidation of the juvenile horseshoe crab to decrease significantly.
表1 不同盐度下中国鲎幼鲎成活率、蜕壳增重率和蜕壳率Tab. 1 Survival rate, molting weight gain rate and molting rate of the juvenile horseshoe crab under different salinity |
| 盐度/‰ | 成活率/% | 蜕壳率/% | 一龄幼鲎平均体质量/mg | 二龄幼鲎平均体质量/mg | 蜕壳增重率/% |
|---|---|---|---|---|---|
| 5 | 74.81±5.01a | 42.17±6.89a | 34.57±1.81 | 49.26±1.59a | 42.58±3.03a |
| 10 | 83.70±2.57ab | 64.70±5.65b | 33.16±2.88 | 53.94±2.35ab | 63.17±2.03bc |
| 15 | 88.15±1.28bc | 75.62±8.82bc | 34.53±2.30 | 55.60±3.87ab | 61.02±2.11bc |
| 20 | 96.30±6.41c | 82.96±7.14c | 35.08±3.21 | 56.62±3.68ab | 61.70±3.71bc |
| 25 | 93.33±2.23bc | 92.08±1.20c | 33.33±1.45 | 57.30±2.29b | 71.92±2.54c |
| 30 | 87.41±4.63bc | 91.46±1.97c | 32.44±2.37 | 55.39±2.05ab | 70.98±3.99c |
| 35 | 85.55±4.01abc | 85.89±1.47c | 33.63±1.59 | 55.20±2.46ab | 64.14±2.46bc |
| 40 | 87.41±1.28bc | 46.69±9.47a | 34.81±0.37 | 53.72±1.54ab | 54.31±1.59b |
注: 同列数据上标字母不同表示存在显著差异(P<0.05), 下表同 |
表2 不同盐度对幼鲎免疫酶活性的影响Tab. 2 Effect of salinity on immune enzyme activity of the juvenile horseshoe crab |
| 盐度/‰ | 一龄幼鲎 | 二龄幼鲎 | ||||
|---|---|---|---|---|---|---|
| LZM/(μg·mL-1) | ACP/(U·gprot-1) | AKP/(U·gprot-1) | LZM/(μg·mL-1) | ACP/(U·gprot-1) | AKP/(U·gprot-1) | |
| 5 | 48.86±1.67 | 9.38±0.45a | 59.14±2.23a | 68.27±9.02 | 7.65±0.43 | 37.30±9.66 |
| 10 | 51.32±5.50 | 9.16±0.11a | 84.39±11.01b | 72.17±13.57 | 7.39±0.37 | 32.93±4.68 |
| 15 | 49.98±10.57 | 9.08±0.32a | 87.02±7.07b | 66.34±11.78 | 7.28±0.19 | 31.52±5.06 |
| 20 | 51.56±1.89 | 9.69±0.38a | 86.72±5.66b | 70.30±7.01 | 7.33±0.31 | 34.37±5.75 |
| 25 | 49.75±4.45 | 9.90±0.94a | 82.83±4.49b | 66.55±4.59 | 7.29±0.16 | 35.53±3.98 |
| 30 | 49.79±4.81 | 9.61±0.32a | 83.67±2.11b | 69.69±4.59 | 7.24±0.37 | 35.63±4.18 |
| 35 | 51.69±1.12 | 12.22±0.96b | 85.56±8.10b | 71.60±4.59 | 7.25±0.28 | 35.94±6.86 |
| 40 | 50.60±2.55 | 12.46±0.96b | 84.85±5.60b | 69.33±4.59 | 7.44±0.18 | 38.11±7.69 |
表3 不同盐度对幼鲎抗氧化酶活性的影响Tab. 3 Effect of salinity on antioxidant enzyme activity of the juvenile horseshoe crab |
| 盐度/‰ | 一龄幼鲎 | 二龄幼鲎 | ||||||
|---|---|---|---|---|---|---|---|---|
| T-AOC /(U·mgprot-1) | GSH-Px /(U·mgprot-1) | CAT /(U·mgprot-1) | SOD /(U·mgprot-1) | T-AOC /(U·mgprot-1) | GSH-Px /(U·mgprot-1) | CAT /(U·mgprot-1) | SOD /(U·mgprot-1) | |
| 5 | 2.03±0.32 | 3.23±1.05 | 17.13±2.11a | 49.30±1.58 | 3.55±0.19 | 8.43±0.64ab | 47.65±8.52 | 143.64±10.01 |
| 10 | 1.70±0.25 | 3.60±1.15 | 17.37±1.68a | 50.54±1.36 | 3.07±0.58 | 7.48±0.74a | 50.73±6.06 | 145.03±15.40 |
| 15 | 1.71±0.19 | 3.10±0.30 | 18.12±1.20a | 50.01±1.51 | 3.08±0.56 | 9.97±0.80b | 54.21±4.54 | 140.72±22.37 |
| 20 | 1.65±0.13 | 3.67±1.22 | 17.48±1.16a | 51.94±2.79 | 2.92±0.45 | 9.60±1.47ab | 49.66±1.91 | 150.22±10.39 |
| 25 | 1.63±0.18 | 3.80±0.50 | 18.20±0.88a | 50.39±2.25 | 2.93±0.43 | 9.87±0.83ab | 49.71±2.29 | 145.14±11.80 |
| 30 | 1.63±0.14 | 3.57±0.68 | 18.40±1.61a | 51.04±1.00 | 2.98±0.78 | 10.07±0.35b | 49.77±8.17 | 145.45±5.25 |
| 35 | 1.86±0.15 | 3.90±0.26 | 20.97±1.705a | 50.42±2.01 | 3.05±0.22 | 9.80±0.76ab | 49.57±3.35 | 143.84±11.49 |
| 40 | 1.95±0.06 | 3.48±1.30 | 27.11±1.78b | 52.00±1.09 | 3.07±0.31 | 10.03±0.91b | 49.48±12.07 | 142.19±10.00 |
| [1] |
鲍虞园, 叶国玲, 颉晓勇, 2020. 中国鲎人工繁育及1龄稚鲎形态性状对体质量的影响[J]. 渔业科学进展, 41(4): 77-84.
|
| [2] |
蔡星媛, 张秀梅, 田璐, 等, 2015. 盐度胁迫对魁蚶稚贝血淋巴渗透压及鳃Na+/K+-ATP酶活力的影响[J]. 南方水产科学, 11(2): 12-19.
|
| [3] |
陈垂坤, 郜卫华, 田罗, 2015. 盐度和营养素对甲壳动物营养生理的影响研究进展[J]. 长江大学学报(自然科学版), 12(3): 47-51. (in Chinese)
|
| [4] |
程鹏, 周爱娜, 霍淑芳, 等, 2006. 中国鲎人工培育的幼体对不同环境适应性的研究[J]. 厦门大学学报(自然科学版), 45(3): 404-408.
|
| [5] |
程文, 2007. 中国鲎的胚胎发育及温度和盐度对一期幼体代谢的影响[D]. 武汉: 华中农业大学: 26-33.
|
| [6] |
高凤英, 廖永岩, 叶富良, 2003. 中国鲎幼体阶段(黄皮鲎)饵料的初步研究[J]. 海洋通报, 22(4): 92-96.
|
| [7] |
洪水根, 2011. 中国鲎生物学研究[M]. 厦门: 厦门大学出版社: 58-62.
|
| [8] |
蒋湘, 谢妙, 彭树锋, 等, 2017. 盐度对日本囊对虾生长与存活率的影响[J]. 江苏农业科学, 45(16): 152-155.
|
| [9] |
李锋, 廖永岩, 董学兴, 1999. 盐度对中国鲎(Tachypleus tridentatus)胚胎发育的影响[J]. 湛江海洋大学学报, 19(3): 4-8.
|
| [10] |
李娜, 王仁杰, 赵玉超, 等, 2017. 高盐胁迫对凡纳滨对虾生长指标、血浆渗透压及Na+-K+-ATP酶活力的影响[J]. 浙江海洋学院学报(自然科学版), 36(3): 196-201.
|
| [11] |
李琼珍, 2010. 中国鲎保育工作研究进展[J]. 生物学杂志, 27(4): 71-74.
|
| [12] |
李琼珍, 胡梦红, 2011. 广西北海中国鲎资源现状以及保育工作进展[J]. 海洋环境科学, 30(1): 131-134.
|
| [13] |
李志辉, 王玉, 李光光, 等, 2019. 温度和盐度对脊尾白虾生长和非特异性免疫的影响[J]. 海洋湖沼通报, (6): 78-82.
|
| [14] |
梁君荣, 王军, 苏永全, 等, 2001. 四种重金属对中国鲎(Tachypleus tridentatus)胚胎发育的影响[J]. 生态学报, 21(6): 1009-1012.
|
| [15] |
柳旭东, 王际英, 张利民, 等, 2009. 影响水产动物Na+-K+-ATP酶活力的水环境因素[J]. 水产科学, 28(3): 171-175.
|
| [16] |
龙晓文, 吴旭干, 刘智俊, 等, 2014. 盐度对脊尾白虾存活、生长和蜕壳的影响[J]. 广东农业科学, 41(23): 111-115, 130.
|
| [17] |
路允良, 王芳, 赵卓英, 等, 2012. 盐度对三疣梭子蟹生长、蜕壳及能量利用的影响[J]. 中国水产科学, 19(2): 237-245.
|
| [18] |
亓磊, 顾孝连, 蒋科技, 等, 2013. 盐度对拟穴青蟹幼蟹存活、生长和Na+/K+-ATP酶活性的影响[J]. 海洋科学, 37(2): 56-60.
|
| [19] |
申玉春, 陈作洲, 刘丽, 等, 2012. 盐度和营养对凡纳滨对虾蜕壳和生长的影响[J]. 水产学报, 36(2): 290-299.
|
| [20] |
王德祥, 苏永全, 王军, 等, 2001. 几种因子对中国鲎胚胎和幼体发育的影响[J]. 中国水产科学, 8(3): 10-14.
|
| [21] |
王军, 王德祥, 苏永全, 等, 2001. 中国鲎的胚胎发育[J]. 动物学杂志, 36(4): 9-14.
|
| [22] |
翁朝红, 谢仰杰, 肖志群, 等, 2012. 福建及中国其他沿岸海域中国鲎资源分布现状调查[J]. 动物学杂志, 47(3): 40-48.
|
| [23] |
吴丹华, 郑萍萍, 张玉玉, 等, 2010. 温度胁迫对三疣梭子蟹血清中非特异性免疫因子的影响[J]. 大连水产学院学报, 25(4): 370-375.
|
| [24] |
颜明艳, 李琼珍, 宋洁, 等, 2019. 基于MAXENT模型评估北部湾潮间带中国鲎和圆尾鲎稚鲎的潜在地理分布及种群保育对策[J]. 生态学报, 39(9): 3100-3109.
|
| [25] |
杨其彬, 叶乐, 温为庚, 等, 2008. 盐度对斑节对虾蜕壳、存活、生长和饲料转化率的影响[J]. 南方水产, 4(1): 16-21.
|
| [26] |
杨宇晴, 余德光, 谢骏, 等, 2010. 急性盐度胁迫对斜带石斑鱼Na+/K+-ATP酶及血清应激指标的影响[J]. 热带海洋学报, 29(4): 160-164.
|
| [27] |
游出超, 董兰芳, 曾梦清, 等, 2019. 急性低盐度胁迫对可口革囊星虫存活、Na+/K+-ATP酶及磷酸酶活力的影响[J]. 海洋科学, 43(3): 82-89.
|
| [28] |
于忠利, 乔振国, 王建纲, 2010. 盐度和水流对青蟹蜕壳及存活的影响[J]. 海洋科学, 34(10): 68-70.
|
| [29] |
赵国庆, 邱盛尧, 黄洪亮, 等, 2018. 盐度骤变和渐变对南极大磷虾生存状况和蜕壳的影响[J]. 海洋渔业, 40(3): 360-367.
|
| [30] |
朱俊华, 吴宙, 冯炳斌, 等, 2020. 全球中华鲎资源保护现状及对策建议[J]. 生物多样性, 28(5): 621-629.
|
| [31] |
|
| [32] |
|
| [33] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}