
罗氏沼虾线粒体锰超氧化物歧化酶的克隆表达与功能研究
|
李亚男(1987—), 男, 河南省南阳市人, 特聘副教授, 从事水生生物抗氧化相关研究。email: |
Copy editor: 林强
收稿日期: 2021-08-26
修回日期: 2021-11-04
网络出版日期: 2021-11-09
基金资助
国家自然科学基金(42006115)
广东省渔业生态环境重点实验室开放课题(FEEL-2021-10)
广东省农业农村厅乡村振兴战略专项(200-2018-XMZC-0001-107-0298)
Molecular cloning and functional study of mitochondrial manganese superoxide dismutase in Macrobrachium rosenbergii
Copy editor: LIN Qiang
Received date: 2021-08-26
Revised date: 2021-11-04
Online published: 2021-11-09
Supported by
National Natural Science Foundation of China(42006115)
Fund of Guangdong Provincial Key Laboratory of Fishery Ecology and Environment(FEEL-2021-10)
Guangdong Provincial Rural Revitalization Strategy Special Fund(200-2018-XMZC-0001-107-0298)
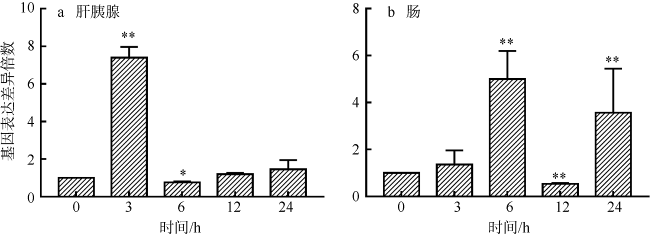
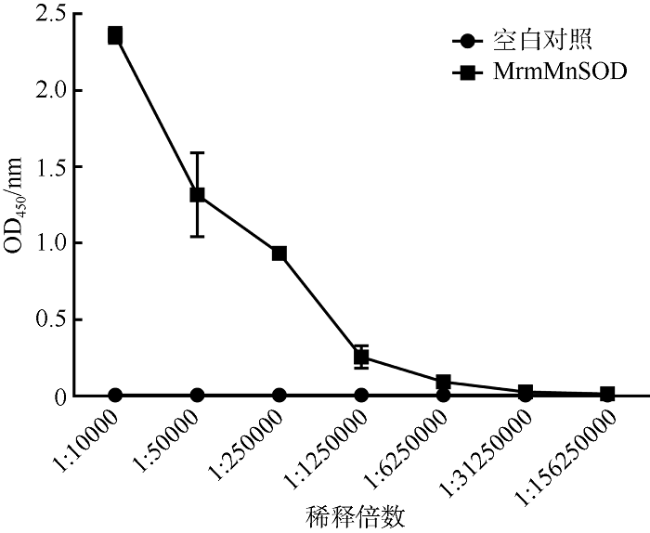
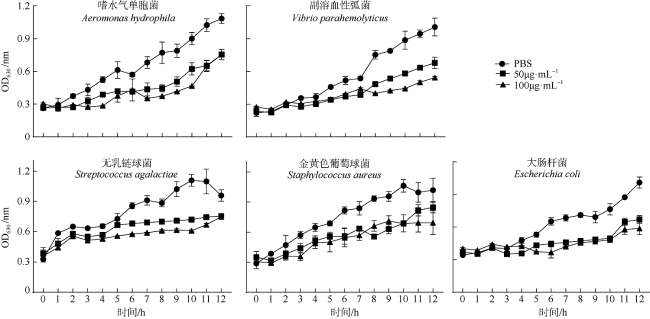
罗氏沼虾(Macrobrachium rosenbergii)是我国重要的经济虾类之一。近年来, 细菌性感染造成罗氏沼虾养殖业病害频发, 经济损失巨大。因此了解其免疫机制对于指导疾病防控至关重要。线粒体锰超氧化物歧化酶(mitochondrial manganese superoxide dismutase, mMnSOD)被认为是抵御氧化应激的第一道防线, 在先天性免疫中具有至关重要的作用。目前, 甲壳动物中mMnSOD的免疫功能尚不清楚。为此, 本研究克隆了罗氏沼虾mMnSOD基因(mMnSOD of M. rosenbergii, MrmMnSOD), 制备了多克隆抗体, 分析了嗜水气单胞菌(Aeromonas hydrophila)感染下, MrmMnSOD在不同组织中的表达模式。结果显示, 肝胰腺和肠组织中MrmMnSOD的基因表达水平分别在感染后3h和6h达到最大; 组织免疫荧光分析显示, 肝胰腺和肠组织中MrmMnSOD均在感染后12h达到最大荧光强度。以上结果表明, mMnSOD参与了罗氏沼虾对嗜水气单胞菌的免疫应激反应。进一步的蛋白抑菌实验表明, MrmMnSOD可显著抑制大肠杆菌(Escherichia coli)、嗜水气单胞菌、副溶血弧菌(Vibrio parahaemolyticus)、金黄色葡萄球菌(Staphylococcus aureus)和无乳链球菌(Streptococcus agalactiae)的生长, 推测该蛋白可能属于免疫相关分子, 可通过抑菌反应发挥免疫功能。当前的研究结果进一步丰富了甲壳动物先天性免疫基础理论, 也可为今后罗氏沼虾的病害防控和药物研发提供新的靶点参考。
李亚男 , 陆霖青 , 张鹏 , 秦真东 , 林蠡 , 晏磊 . 罗氏沼虾线粒体锰超氧化物歧化酶的克隆表达与功能研究[J]. 热带海洋学报, 2022 , 41(3) : 111 -118 . DOI: 10.11978/2021109
Macrobrachium rosenbergii is one of the important shrimps for consumers in China. In recent years, bacterial diseases have occurred frequently and caused huge economic losses in M. rosenbergii aquaculture. Therefore, understanding its immune mechanism is essential to guide disease prevention and control. Mitochondrial manganese superoxide dismutase (mMnSOD) is considered to be the first line against oxidative stress, playing a vital role in innate immunity. However, the immune function of mMnSOD is still unclear. To this end, we cloned the mMnSOD of M. rosenbergii (MrmMnSOD), prepared its polyclonal antibodies, and analyzed its expression patterns under Aeromonas hydrophila infection. On the gene expression levels, MrmMnSOD reached a peak after 3- and 6-h infection in hepatopancreas and intestine tissue, respectively. Tissue immunofluorescence analysis showed that the maximum fluorescence intensity occurred at 12 h after infections in both hepatopancreas and intestine tissue. The above results indicated that MrmMnSOD is involved in the immune response against A. hydrophila. Further antibacterial assays showed that MrmMnSOD significantly inhibited the growth of Escherichia coli, A. hydrophila, Vibrio parahaemolyticus, Staphylococcus aureus, and Streptococcus agalactiae, suggesting that MrmMnSOD may belong to an immune-related molecule, which exerts immune function by antibacterial effect. These results enrich the basic theory in crustacean innate immunity, and provide a new target for disease control and drug research for M. rosenbergii breeding in the future.
Key words: Macrobrachium rosenbergii; SOD; expression pattern; antibacterial assay
表1 引物序列Tab. 1 Primers used in the study |
| 引物 | 序列 (5′—3′) | 用途 |
|---|---|---|
| MrmMnSOD S1 | CCCTGAAGTTCAATGGAGGAGGT | qRT-PCR |
| MrmMnSOD A1 | TTGGGACAAGTAGCAATTTGTAGTG | |
| Mr18S F | TAGCAATTCGCCGTCGTTATTC | qRT-PCR内参 |
| Mr18S R | CTACCCCCGGAACTCAAAGACT | |
| MrmMnSOD-32a S | TATTTTCAGGGATCCGAATTCATGCTCAGTTTCAGCCGTCTG | |
| MrmMnSOD-32a A | GTGGTGGTGGTGGTGCTCGAGTCACTTTGCAGCAGCAAATCTT | 载体构建 |
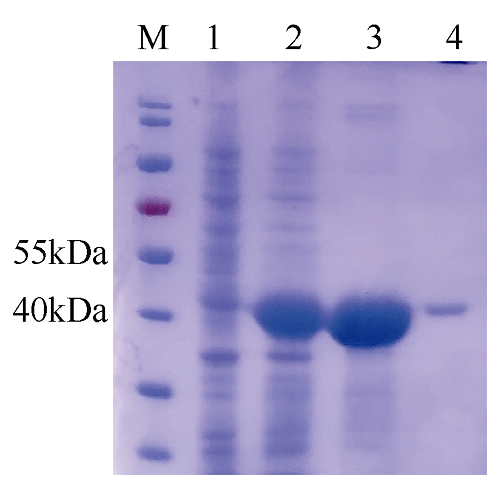
图2 罗氏沼虾mMnSOD的SDS-PAGE检测图M: 蛋白marker, 1: IPTG诱导前菌体总蛋白, 2: IPTG诱导后菌体总蛋白, 3: 诱导后包涵体总蛋白, 4: 纯化后的重组蛋白 Fig. 2 SDS-PAGE detection of M. rosenbergii mitochondrial manganese SOD M, protein marker; 1: total protein from E. coli BL21 (DE3) before IPTG induction; 2: total protein from E. coli BL21 (DE3) after IPTG induction; 3: total protein of the inclusion body; 4: the purified recombinant protein |
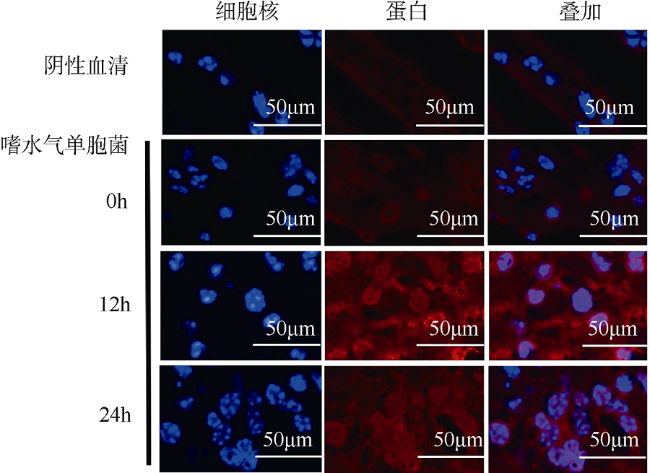
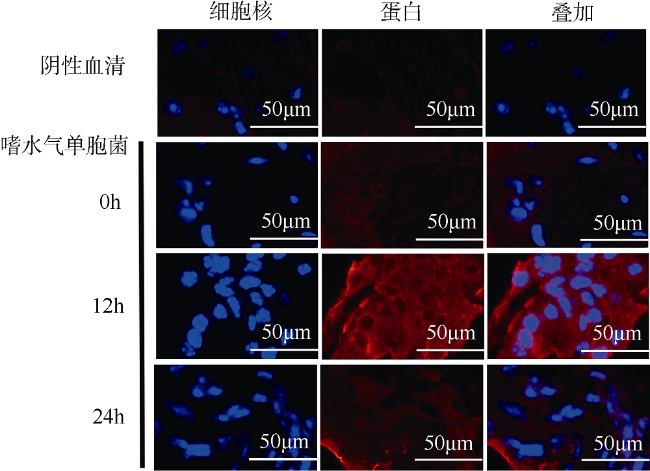
图4 嗜水气单胞菌(A. hydrophila)感染后肝胰腺组织免疫荧光分析红色荧光标记MrmMnSOD蛋白, 蓝色荧光标记细胞核 Fig. 4 Immunofluorescence analysis of hepatopancreas after A. hydrophila infection. Red fluorescence represents MrmMnSOD; and blue, nucleus |
| [1] |
董学洪, 陈正兴, 2015. 罗氏沼虾养殖中常见病害及防治[J]. 水产养殖, 36(11): 46-48. (in Chinese)
|
| [2] |
李亚男, 张海滨, 2018. 海洋无脊椎动物抗氧化酶研究进展[J]. 海洋通报, 37(3): 241-253.
|
| [3] |
肖楚康, 方刘, 阮国良, 等, 2019. 罗氏沼虾淡化养殖的现状与展望[J]. 江苏农业科学, 47(8): 29-33.
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}