
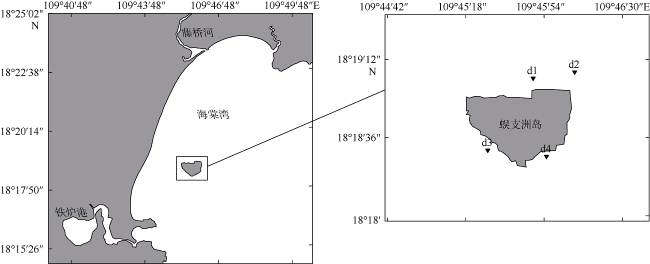
三亚蜈支洲岛海洋牧场近岛区底表大型底栖动物群落结构及评价
|
马文刚(1997—), 男, 山东省济南市人, 硕士研究生, 从事群落生态学研究。email: |
Copy editor: 林强
收稿日期: 2021-09-15
修回日期: 2021-11-23
网络出版日期: 2021-11-29
基金资助
国家重点研发计划课题(2019YFD0901304)
国家自然科学基金(42076097)
海南省自然科学基金(2019RC070)
Community structure evaluation of epifaunal macrozoobenthos in the near-island waters of marine ranching in Wuzhizhou Island, Sanya
Copy editor: LIN Qiang
Received date: 2021-09-15
Revised date: 2021-11-23
Online published: 2021-11-29
Supported by
National Key R&D Program of China(2019YFD0901304)
National Natural Science Foundation of China(42076097)
Hainan Natural Science Foundation(2019RC070)
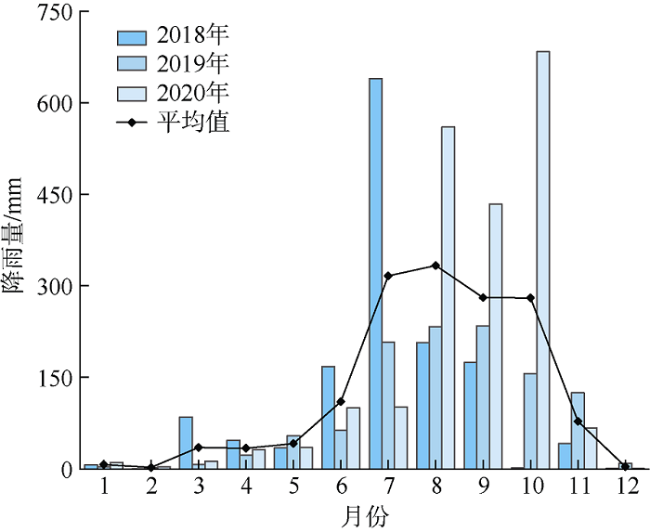
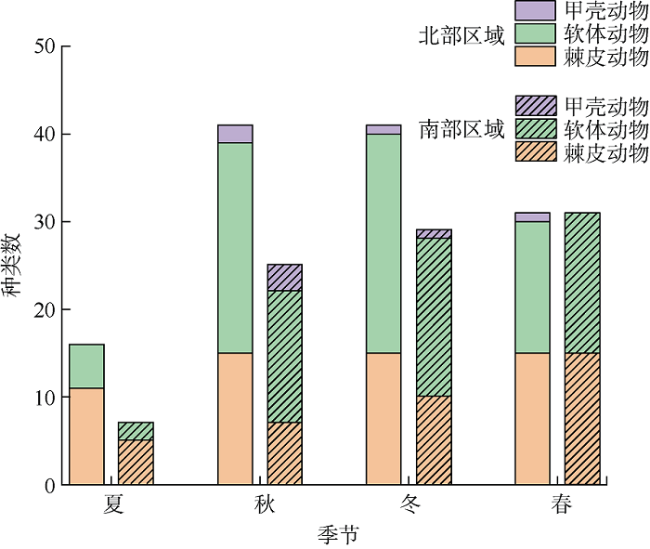
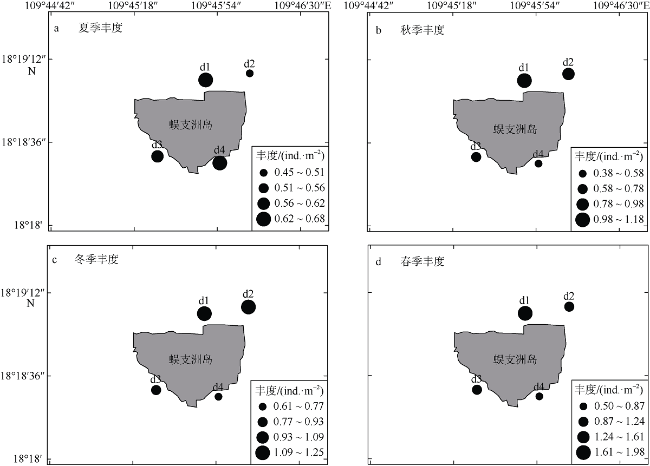
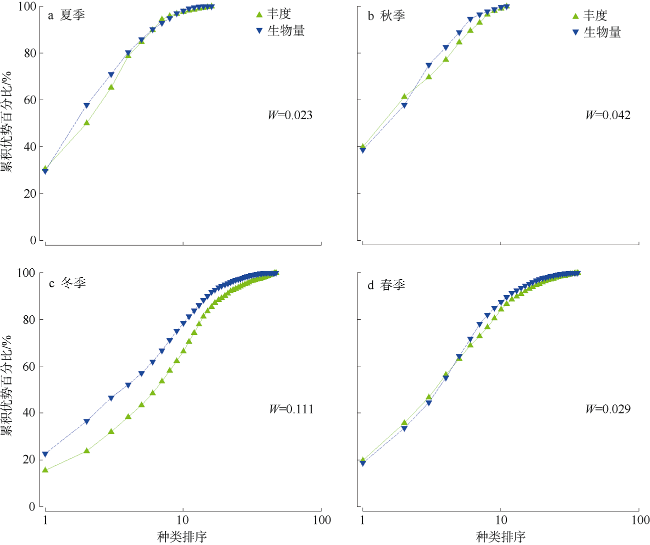
三亚蜈支洲岛海洋牧场拥有丰富的岛礁生物资源。为掌握海洋牧场近岛区的底表大型底栖动物群落组成和分布特征及其影响因子、评估海洋牧场底栖生态系统健康状况, 本研究于2020—2021年进行了底表大型无脊椎动物群落季节变动的调查。结果表明: 近岛珊瑚礁区共鉴定出棘皮动物、软体动物、节肢动物3大门类90种, 其中秋季种类数最多为55种, 夏季种类数最少为16种; 底表大型底栖动物的年平均丰度为0.87±0.26ind.·m-2, 年平均生物量为76.99±34.32g·m-2。群落聚类分析(cluster)与多维排序尺度分析(multidimensional Scaling, MDS)表明, 该区域群落结构季节性差异不显著, 各站位间群落结构受沉积物性质以及人类活动的频繁程度的影响, 形成北部与南部区域2个聚类组。全年的物种丰富度指数d为2.7±1.16, 多样性指数H′为3.14±0.88, 均匀度指数J为0.76±0.11。基于多样性指数H'及多变量海洋生物指数(Multivariate-AZTI′s marine biotic index, M-AMBI)评价指标, 海洋牧场近岛区环境除夏季为轻度污染外, 其他季节均为无污染状态。采用动物丰度与生物量比较曲线(abundance-biomass curves, ABC曲线)法评价底栖动物群落稳定性状况得出, 除冬季以外, 其他季节底表大型底栖动物群落受到一定程度干扰, 尤其是夏季群落结构稳定性较低。建议应持续关注底表大型底栖动物群落变动, 调整、优化涉海休闲旅游活动, 以保证蜈支洲岛海洋牧场生态系统的长期健康与稳定。
马文刚 , 夏景全 , 魏一凡 , 尹洪洋 , 覃乐政 , 刘相波 , 胡雪晴 , 许强 , 李秀保 , 王爱民 . 三亚蜈支洲岛海洋牧场近岛区底表大型底栖动物群落结构及评价[J]. 热带海洋学报, 2022 , 41(3) : 135 -146 . DOI: 10.11978/2021125
Marine ranching in Wuzhizhou Island was rich in reef biological resources. The composition, distribution characteristics, and influencing factors of epifaunal macrozoobenthos community in the near-island area of marine ranching were studied; and the health status of marine ranching ecosystem was evaluated, based on samples obtained from the epibenthic macroinvertebrates community during 2020-2021. Ninety species of echinoderms, molluscs, and arthropods were identified in the near-island coral reef area, of which 55 species were the most in autumn and 16 species were the least in summer. The average annual abundance of epifaunal macrozoobenthos was 0.87±0.26 ind.·m-2, and the average annual biomass was 76.99±34.32 g·m-2. Community cluster analysis and Multi-dimensional scaling analysis showed that the seasonal difference of community structure in this area was not significant. The community structure among stations was affected by the nature of sediments and the frequency of human activities, forming two cluster groups in the north and south areas. The Margalef's richness index (d) was 2.7±1.16, Shannon-Wiener index (H') was 3.14±0.88, and Pielou's evenness index (J) was 0.76±0.11. Based on the diversity evaluation index and Multivariate-AZTI′s marine biotic index (M-AMBI), the environment of marine ranching near the island was not polluted in most seasons except for summer when it was lightly polluted. The stability of macrozoobenthos community was evaluated by abundance-biomass curves. We conclude that the epifaunal macrozoobenthos community was disturbed to a certain extent in all seasons except winter, especially in summer. We should continue to pay attention to the changes of epifaunal macrozoobenthos community and adjust recreational tourism activities so as to ensure the long-term health and stability of marine ranching ecosystem in Wuzhizhou Island.
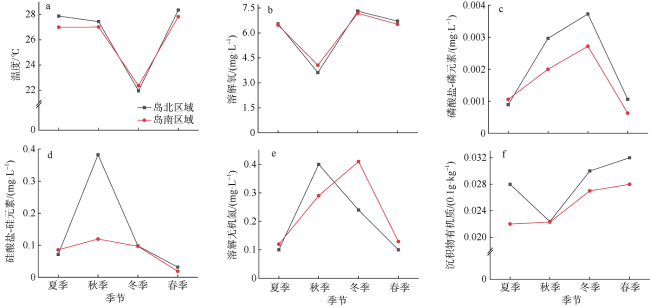
表1 近岛区水体环境因子季节变化Tab. 1 Seasonal variation of water environmental factors in coastal area of Wuzhizhou Island |
| 季节 | T/℃ | DO/(mg·L-1) | Chl a/(μg·L-1) | P/(mg·L-1) | Si/(mg·L-1) | DIN/(mg·L-1) | Md | Org/(0.1g·kg-1) |
|---|---|---|---|---|---|---|---|---|
| 夏季 | 27.44±0.70ab | 6.51±0.07b | 0.37±0.08ab | 0.001±0.0002b | 0.075±0.011ab | 0.112±0.011b | 1.89±1.31a | 2.60±0.94a |
| 秋季 | 27.24 ±0.30b | 3.84±0.68c | 2.03±0.34a | 0.003±0.0009ab | 0.379±0.278a | 0.347±0.068a | — | 2.23±0.28a |
| 冬季 | 22.16±0.24c | 7.25±0.08a | 0.49±0.29b | 0.003±0.0014a | 0.095±0.013ab | 0.324±0.120a | 2.26±2.47a | 2.87±0.43a |
| 春季 | 28.10±0.31a | 6.62±0.17b | 0.57±0.14ab | 0.001±0.0005b | 0.030±0.013b | 0.112±0.027b | 2.37±1.48a | 3.10±0.36a |
注: “—”表示数据缺失; 同一列中不同字母上标表示差异显著(P<0.05), 相同字母表示差异不显著(P>0.05)。T: 温度; DO: 溶解氧; Chl a: 叶绿素a; P: 磷酸盐-磷元素; Si: 硅酸盐-硅元素; DIN: 溶解无机氮; Md: 中值粒径; Org: 沉积物有机质 |
表2 各季节优势种Tab. 2 Dominant species in each season |
| 优势种 | 优势度 | ||||
|---|---|---|---|---|---|
| 种名 | 学名 | 夏季 | 秋季 | 冬季 | 春季 |
| 吕宋棘海星 | Echinaster luzonicus | 0.046 | 0.042 | 0.038 | 0.037 |
| 蓝指海星 | Linckia laevigata | 0.060 | 0.032 | 0.043 | 0.039 |
| 蓝环冠海胆 | Diadema savignyi | — | — | 0.038 | 0.078 |
| 刺冠海胆 | Diadema setosum | — | 0.049 | — | — |
| 冠刺棘海胆 | Echinothrix diadema | — | — | 0.029 | — |
| 红腹海参 | Holothuria eduils | 0.114 | 0.225 | 0.057 | 0.191 |
| 黑海参 | Holothuria atra | 0.305 | 0.121 | 0.060 | 0.106 |
| 绿刺参 | Stichopus chloronotus | 0.101 | — | 0.034 | 0.055 |
| 金口蝾螺 | Turbo argyrostomus | 0.050 | 0.028 | 0.047 | 0.028 |
| 塔形扭柱螺 | Tectus pyramis | — | — | 0.038 | — |
| 角小核果螺 | Drupella cornus | 0.146 | — | 0.107 | 0.028 |
| 紫栖珊瑚螺 | Coralliophila violacea | — | — | 0.023 | — |
| 焦棘螺 | Chicoreus torrefactus | — | — | 0.036 | — |
| 秉氏海齿花 | Comanthus bennetti | — | 0.048 | 0.047 | 0.094 |
| 许氏大羽花 | Comanthina schlegeli | — | — | 0.023 | 0.049 |
| 小足真寄居蟹 | Dardanus pedunculatus | — | 0.084 | — | — |
注: “—”表示非优势种或调查中未发现 |
表3 各季节生物多样性指数及M-AMBI指数值Tab. 3 Biodiversity index and M-AMBI in four seasons |
| d | J | H′ | 水体状况 | M-AMBI | 水体环境状况 | |
|---|---|---|---|---|---|---|
| 夏季 | 1.26±0.53b | 0.70±0.10a | 2.14±0.41b | 轻度污染 | 0.59±0.06b | 优良 |
| 秋季 | 3.11±1.24ab | 0.74±0.13a | 3.21±0.93a | 无污染 | 0.77±0.16a | 高等 |
| 冬季 | 3.40±0.26a | 0.81±0.09a | 3.71±0.52a | 无污染 | 0.85±0.07a | 高等 |
| 春季 | 3.04±0.99ab | 0.79±0.13a | 3.52±0.75a | 无污染 | 0.82±0.09a | 高等 |
注: 同一列中不同字母上标表示差异显著(P<0.05), 相同字母表示差异不显著(P>0.05) |
| [1] |
蔡立哲, 马丽, 高阳, 等, 2002. 海洋底栖动物多样性指数污染程度评价标准的分析[J]. 厦门大学学报(自然科学版), 41(5): 641-646.
|
| [2] |
陈晨, 焦海峰, 王一农, 等, 2016. 象山港海洋牧场示范区大型底栖生物的时空变化[J]. 海洋与湖沼, 47(1): 130-139.
|
| [3] |
董栋, 李新正, 王洪法, 等, 2015. 海南岛三亚珊瑚礁区大型底栖动物群落特征[J]. 海洋科学, 39(3): 83-91.
|
| [4] |
黄丁勇, 王建佳, 陈甘霖, 等, 2021. 亚龙湾珊瑚礁大型礁栖生物的群落结构及生态警示[J]. 生态学杂志, 40(2): 412-426.
|
| [5] |
黄端杰, 许强, 李秀保, 等, 2020. 三亚蜈支洲岛珊瑚礁-沙质底复合区棘皮动物群落结构[J]. 海洋与湖沼, 51(1): 103-113.
|
| [6] |
黄晖, 张浴阳, 刘骋跃, 2020. 热带岛礁型海洋牧场中珊瑚礁生境与资源的修复[J]. 科技促进发展, 16(2): 225-230.
|
| [7] |
李秀保, 李元超, 许强, 2019. 三亚蜈支洲岛珊瑚礁的现状、生态修复及保护对策[M]. 北京: 科学出版社: 14-84. (in Chinese)
|
| [8] |
李元超, 杨毅, 郑新庆, 等, 2015. 海南三亚后海海域珊瑚礁生态系统的健康状况及其影响因素[J]. 生态学杂志, 34(4): 1105-1112.
|
| [9] |
林和山, 俞炜炜, 刘坤, 等, 2015. 基于AMBI和M-AMBI法的底栖生态环境质量评价-以厦门五缘湾海域为例[J]. 海洋学报, 37(8): 76-87.
|
| [10] |
刘帅磊, 王赛, 崔永德, 等, 2018. 亚热带城市河流底栖动物完整性评价--以流溪河为例[J]. 生态学报, 38(1): 342-357.
|
| [11] |
罗章凤, 方展强, 2017. 珠海横琴岛芒洲湿地红树林人工恢复期大型底栖动物群落结构研究[J]. 热带海洋学报, 36(3): 61-72.
|
| [12] |
全秋梅, 肖雅元, 徐姗楠, 等, 2020. 胶州湾大型底栖动物群落结构季节变化及其与环境因子的关系[J]. 生态学杂志, 39(12): 4110-4120.
|
| [13] |
孙有方, 雷新明, 练健生, 等, 2018. 三亚珊瑚礁保护区珊瑚礁生态系统现状及其健康状况评价[J]. 生物多样性, 26(3): 258-265.
|
| [14] |
田胜艳, 于子山, 刘晓收, 等, 2006. 丰度/生物量比较曲线法监测大型底栖动物群落受污染扰动的研究[J]. 海洋通报, 25(1): 92-96.
|
| [15] |
田胜艳, 张文亮, 张锐, 2009. 大型底栖动物在海洋生态系统中的作用[J]. 盐业与化工, 38(2): 50-54.
|
| [16] |
王言丰, 胡启伟, 余景, 等, 2019. 粤东柘林湾海洋牧场渔业资源增殖效果评估[J]. 南方水产科学, 15(2): 12-19.
|
| [17] |
吴莹莹, 雷新明, 黄晖, 等, 2021. 南海典型珊瑚礁生态系统健康评价方法研究[J]. 热带海洋学报, 40(4): 84-97.
|
| [18] |
吴钟解, 张光星, 陈石泉, 等, 2015. 海南西瑁洲岛周边海域造礁石珊瑚空间分布及其生态系统健康评价[J]. 应用海洋学学报, 34(1): 133-140.
|
| [19] |
夏景全, 贾志宇, 张国豪, 等, 2020. 火山石对破碎化珊瑚礁的修复效果研究[J]. 浙江海洋大学学报(自然科学版), 39(3): 237-244.
|
| [20] |
夏景全, 2021. 火山石生境重构与珊瑚移植在修复三亚破碎化珊瑚礁上的作用[D]. 海南: 海南大学.
|
| [21] |
许惠丽, 冯博轩, 谢敏睿, 等, 2020. 三亚蜈支洲岛两种造礁石珊瑚的生理特征[J]. 应用海洋学学报, 39(2): 181-188.
|
| [22] |
许强, 刘维, 高菲, 等, 2018. 发展中国南海热带岛礁海洋牧场--机遇、现状与展望[J]. 渔业科学进展, 39(5): 173-180.
|
| [23] |
徐帅帅, 邸宝平, 王玉珏, 等, 2017. 我国典型潮间带底栖硅藻群落空间分布特征[J]. 海洋学报, 39(6): 95-113.
|
| [24] |
徐兆礼, 陈亚瞿, 1989. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J]. 生态学杂志, 8(4): 13-15, 19.
|
| [25] |
杨东, 周政权, 张建设, 等, 2017. 烟台牟平海洋牧场夏季大型底栖动物群落特征[J]. 海洋科学, 41(5): 134-143.
|
| [26] |
袁涛萍, 李恒翔, 李路, 等, 2017. 夏季大亚湾大型底栖动物群落结构[J]. 热带海洋学报, 36(1): 41-47.
|
| [27] |
张志南, 2000. 水层-底栖耦合生态动力学研究的某些进展[J]. 青岛海洋大学学报, 30(1): 115-122.
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}