
两种蓝细菌中7对毒素-抗毒素系统的分析和鉴定
|
林世团(1995—), 男, 广东省揭阳市人, 硕士, 从事毒素-抗毒素系统的功能研究。email: |
Copy editor: 林强
收稿日期: 2021-10-26
修回日期: 2021-12-01
网络出版日期: 2021-12-10
基金资助
国家自然科学基金(31625001)
国家自然科学基金(91951203)
Analysis and identification of seven toxin-antitoxin systems in two cyanobacteria
Copy editor: LIN Qiang
Received date: 2021-10-26
Revised date: 2021-12-01
Online published: 2021-12-10
Supported by
National Natural Science Foundation of China(31625001)
National Natural Science Foundation of China(91951203)
蓝细菌(cyanobacteria)是一类能进行放氧光合作用的原核生物。蓝细菌生长速度较快, 几乎存在于所有的陆地和水生环境中。毒素-抗毒素(toxin-antitoxin, TA)系统在原核生物中分布十分广泛, 在细菌的生命活动中扮演了重要的角色, 如维持水平基因转移元件的稳定性以及应对环境胁迫压力等。已有的基因组分析表明, 蓝细菌基因组中含有大量潜在的毒素-抗毒素系统, 但是目前对蓝细菌毒素-抗毒素系统的实验鉴定仍较少。本文分别以淡水和海水代表菌株集胞藻PCC6803(Synechocystis sp. PCC6803)和聚球藻WH7803(Synechococcus sp. WH7803)为研究对象, 对二者基因组上的毒素-抗毒素系统进行预测, 并选取预测结果中的7对潜在的毒素-抗毒素系统进行实验验证。结果表明, 集胞藻PCC6803的两对毒素-抗毒素系统BAD01932-1933 和 BAA18559-New ORF7中的毒素具有明显的细胞毒性。本文的研究结果有助于后续对蓝细菌毒素-抗毒素系统及其生态和生物学功能的研究。
关键词: 毒素-抗毒素系统; 蓝细菌; 集胞藻 PCC6803; 聚球藻 WH7803
林世团 , 王晓雪 , 陈冉 . 两种蓝细菌中7对毒素-抗毒素系统的分析和鉴定[J]. 热带海洋学报, 2022 , 41(3) : 119 -134 . DOI: 10.11978/2021144
Cyanobacteria are the only oxygenic photosynthetic prokaryotes on Earth, possessing a relatively fast growth rate and prospering in a range of diverse habitats. Toxin-antitoxin (TA) systems are prevalent in prokaryotes, and play important roles in cell physiological activities including maintenance of mobile genetic elements and mediation of stress responses. Previous bioinformatic studies revealed that putative TA systems are widely distributed in the cyanobacteria genomes. However, studies of TA systems in cyanobacteria are limited. In this study, we predicted TA systems in the genomes of Synechocystis sp. PCC6803 and Synechococcus sp. WH7803, the model strains of freshwater and marine environments cyanobacteria, respectively. The toxicity test of five TA systems in PCC6803 and two TA systems in WH7803 were further implemented. We found that the ectopic expression of toxin New ORF7 and BAD01932 lead to growth inhibition in Escherichia coli host. Our study provides vital information for future study of the ecological and biological roles of TA systems in cyanobacteria.
附表1 本文所用引物Tab. S1 Primers used in this study |
| 引物名称 | 序列(5'—3') | 描述 |
|---|---|---|
| pBAD-01933-F | GGGCTAACAGGAGGAATTAACATGACCCAGATCACCAAGTCCA | 过表达 |
| pBAD-01933-R | GTTTTTGTTCTACGTAAGCTTTCAATTTTTAACACCATCAATCTCTATT | 过表达 |
| pBAD-01932-F | GGGCTAACAGGAGGAATTAACATGGTGTTAAAAATTGAACTTCTTGATA | 过表达 |
| pBAD-01932-R | GTTTTTGTTCTACGTAAGCTTCTAAAAGAGTCTTTCCACCGATTTC | 过表达 |
| Pet28apsp-01933-F | GGGCTAACAGGAGGAATTAACATG GTGTTAAAAATTGAACTTCT | 蛋白表达 |
| Pet28apsp-01933-R | GGTGGTGGTGGTGGTGCTCGAG CTAAAAGAGTCTTTCCACCG | 蛋白表达 |
| Pet28apsp-01932-F | GGGCTAACAGGAGGAATTAACATG ACCCAGATCACCAAGTCCAA | 蛋白表达 |
| Pet28apsp-01932-R | GGTGGTGGTGGTGGTGCTCGAG TCAATTTTTAACACCATCAA | 蛋白表达 |
| pBAD-New ORF7-F | ACAGGAGGAATTAACCATGGAT ATGTTACCTGAAATTAATATTT | 过表达 |
| pBAD-New ORF7-R | GTTTTTGTTCTACGTAAGCTT CTATTTTTTCTCGATAATTTCA | 过表达 |
| pBAD-BAA18559-F | ACAGGAGGAATTAACCATGGAT ATGAATTATCCAATTGTTATTT | 过表达 |
| pBAD-BAA18559-R | GTTTTTGTTCTACGTAAGCTT CTATGCGGCACTCAACATCTTA | 过表达 |
| Pet28apsp-New ORF7-NdeI-F | TCTGTTCCAGGGGCCCCATATG TTACCTGAAATTAATATTTGCC | 蛋白表达 |
| Pet28apsp-New ORF7-NdeI-R | GGTGGTGGTGGTGGTGCTCGAG CTATGCGGCACTCAACATCTTA | 蛋白表达 |
| Pet28apsp-BAA18559-F | TTAAGAAGGAGATATACATATG TTACCTGAAATTAATATTTGCC | 蛋白表达 |
| Pet28apsp-BAA18559-R | GGTGGTGGTGGTGGTGCTCGAG TGCGGCACTCAACATCTTAATT | 蛋白表达 |
| pBAD-01997-F | GGGCTAACAGGAGGAATTAACATGACATCTGTTTTTCACGTTCTAATC | 过表达 |
| pBAD-01997-R | GTTTTTGTTCTACGTAAGCTTTTAGTTTTTCCACTGACTTCTTAGCTG | 过表达 |
| pBAD-01998-F | GGGCTAACAGGAGGAATTAACATGAGATATAAAAATAATTGTCGAATCAAT | 过表达 |
| pBAD-01998-R | GTTTTTGTTCTACGTAAGCTTTTATCTTAAAACCTGTAGATGAAGTTCTTC | 过表达 |
| pBAD-01871-F | TAACAGGAGGAATTAACATGCGAAGACGCAGACCTACC | 过表达 |
| pBAD-01871-R | GTTTTTGTTCTACGTAAGCTTCTAGACATGCTCAAACTTAACGTATTTG | 过表达 |
| pBAD-01872 -F | TAACAGGAGGAATTAACATGTCTAGCACCCTGGCAATCA | 过表达 |
| pBAD-01872 -R | GTTTTTGTTCTACGTAAGCTTTTAAATCCGCCAGACCAAAGC | 过表达 |
| pBAD-BAA17887-1-58-F | GGGCTAACAGGAGGAATTAACATGAAGGTTAAAGTAATCCTCAAAATCT | 过表达 |
| pBAD-BAA17887-1-58-R | GTTTTTGTTCTACGTAAGCTTTCAAGATTGCTTGAGAATACTGGC | 过表达 |
| pBAD-BAA17887-59-136-F | GGGCTAACAGGAGGAATTAACATGTCTCAAGCAATCAGGTATTACTTTA | 过表达 |
| pBAD-BAA17887-59-136-R | GTTTTTGTTCTACGTAAGCTTTTAAGCCGCAACTGTTACGTAGG | 过表达 |
| pBAD-SynWH7803_1122- F | GGGCTAACAGGAGGAATTAACATGGTGGCGGCTGATCATC | 过表达 |
| pBAD-SynWH7803_1122- R | GTTTTTGTTCTACGTAAGCTTTCACAGCGGAACGGCCTG | 过表达 |
| pBAD-SynWH7803_1121- F | GGGCTAACAGGAGGAATTAACATGACCCATCTCAAGCCATCC | 过表达 |
| pBAD-SynWH7803_1121- R | GTTTTTGTTCTACGTAAGCTTCACCACAGCTGCAGTTTCAGG | 过表达 |
| pBAD-SynWH7803_1701 -F | GGGCTAACAGGAGGAATTAACATGATCCCTGTTCTCTCTGGGTT | 过表达 |
| pBAD-SynWH7803_1701- R | GTTTTTGTTCTACGTAAGCTTTCAACCTGTTGCAGTTGATCGG | 过表达 |
| pBAD-SynWH7803_1700- F | GGGCTAACAGGAGGAATTAACATGGGATTTCAGGCTGTGCC | 过表达 |
| pBAD-SynWH7803_1700- R | GTTTTTGTTCTACGTAAGCTTTCAGGAATGCGGCGGTTG | 过表达 |
| pBAD-01933-F | GGGCTAACAGGAGGAATTAACATGACCCAGATCACCAAGTCCA | 过表达 |
| pBAD-01933-R | GTTTTTGTTCTACGTAAGCTTTCAATTTTTAACACCATCAATCTCTATT | 过表达 |
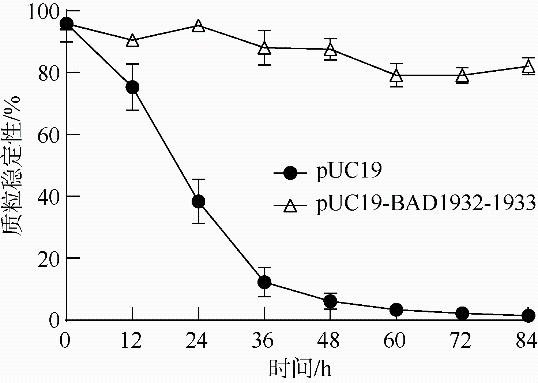
| pUC19-pro-BAD1932-1933-f | GAAAAAGAAGAATTGATGATGT | 质粒稳定性 |
| pUC19-pro-BAD1932-1933-r | CTAAAAGAGTCTTTCCACCGAT | 质粒稳定性 |
| M13-f | CGCCAGGGTTTTCCCAGTCACGAC | 质粒稳定性 |
| M13-r | AGCGGATAACAATTTCACACAGGA | 质粒稳定性 |
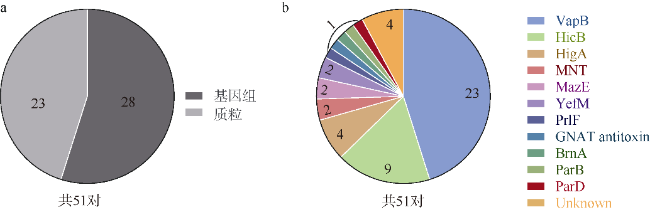
图1 集胞藻PCC6803预测的不同TA类型分析Fig. 1 Analysis of different TA types predicted in Synechocystis sp. PCC6803 |
附表2 集胞藻. PCC6803 毒素-抗毒素系统预测信息Tab. S2 Prediction information of toxin-antitoxin systems in Synechocystis sp. PCC6803 |
| 起始位点 | 终止位点 | 蛋白长度(氨基酸) | 正负链 | 基因功能描述 | 蛋白家族编号 | 蛋白家族描述 | xtalpred 软件分析结果 | 毒素/抗毒素 | |
|---|---|---|---|---|---|---|---|---|---|
| BAA10220 | 2516150 | 2516527 | 125 | - | 未知蛋白 Cag_0280 | PF01850 | PIN 结构域 | VpaC | 毒素 |
| New ORF1 | 2516528 | 2516749 | 73 | - | New ORF | / | / | VapB | 抗毒素 |
| BAA10330 | 2631207 | 2631608 | 133 | - | 毒力相关蛋白毒力相关蛋白C | PF01850 | PIN 结构域 | VpaC | 毒素 |
| BAA10331 | 2631611 | 2631868 | 85 | - | PEMI-样蛋白 | / | / | VapB | 抗毒素 |
| BAA10426 | 2738712 | 2738915 | 67 | - | 未知蛋白 AM1_C0132 | / | / | HicB | 抗毒素 |
| New ORF2 | 2738918 | 2739133 | 71 | - | New ORF | / | / | HicA | 毒素 |
| BAA10766 | 3121193 | 3121687 | 164 | - | 未知蛋白 sll0658 | PF11848 | 未知功能结构域 (DUF3368) | VpaC | 毒素 |
| BAA10767 | 3121684 | 3121953 | 89 | - | 未知蛋白 ssl1255 | PF03683 | 未表征蛋白家族 (UPF0175) | VapB | 抗毒素 |
| BAA10878 | 3262064 | 3262459 | 131 | - | 未知蛋白 sll0525 | PF01850 | PIN 结构域 | VpaC | 毒素 |
| BAA10879 | 3262471 | 3262749 | 92 | - | 未知蛋白 ssl1004 | / | / | VapB | 抗毒素 |
| BAA16615 | 48067 | 48408 | 113 | - | 未知蛋白 sll1400 | PF18480 | 未知功能结构域 (DUF5615) | VpaC | 毒素 |
| BAA16616 | 48405 | 48635 | 76 | - | 未知蛋白 ssl2733 | PF04255 | 未知功能蛋白 (DUF433) | VapB | 抗毒素 |
| BAA16671 | 110392 | 110724 | 110 | + | PrlF | PF15937 | 毒素蛋白YhaV的抗毒素PrlF _ | PrlF_抗毒素 | 抗毒素 |
| BAA16672 | 110808 | 111224 | 138 | + | 未知蛋白 slr0725 | PF11663 | 具有核酸内切酶活性的毒素 | YhaV | 毒素 |
| BAA16930 | 382392 | 382646 | 84 | + | 未知蛋白 ssr1765 | PF15919 | HicB样抗毒素 | HicB | 抗毒素 |
| BAA16931 | 382643 | 382873 | 76 | + | 未知蛋白 ssr1766 | PF07927 | HicA | HicA | 毒素 |
| BAA17012 | 473953 | 474204 | 83 | - | 毒力相关蛋白 C | PF01850 | PIN结构域 | VpaC | 毒素 |
| BAA17013 | 474205 | 474432 | 75 | - | 毒力相关蛋白 B | / | / | VapB | 抗毒素 |
| New ORF3 | 474504 | 474785 | 93 | - | New ORF | / | / | HigB_毒素 | 毒素 |
| BAA17014 | 474789 | 475007 | 72 | - | 未知蛋白 MAE_61720 | / | / | HigA_抗毒素 | 抗毒素 |
| BAA17015 | 475110 | 475445 | 111 | - | 未知蛋白 sll1505 | PF01934 | 未知功能蛋白 DUF86 | HEPN | 毒素 |
| BAA17016 | 475435 | 475743 | 102 | - | 未知蛋白 sll1504 | PF01909 | 核苷酸转移酶结构域 | MNT | 抗毒素 |
| BAA17065 | 545029 | 545253 | 74 | + | 未知蛋白 SYNGTS_0489 | PF04255 | 未知功能蛋白 (DUF433) | VapB | 抗毒素 |
| BAA17066 | 545250 | 545588 | 112 | + | 未知蛋白 SYNGTS_0490 | PF18480 | 未知功能结构域 (DUF5615) | VapC | 毒素 |
| BAA17191 | 679690 | 680178 | 162 | + | 未知蛋白 Ppha_2119 | PF01850 | PIN结构域 | VapC | 毒素 |
| BAA17192 | 680096 | 680368 | 90 | + | 未知蛋白 Ppha_2120 | / | / | VapB | 抗毒素 |
| BAA17278 | 775687 | 776061 | 124 | - | 未知蛋白 sll1912 | PF13470 | PIN结构域 | VapC | 毒素 |
| BAA17279 | 776058 | 776342 | 94 | - | 未知蛋白 ssl3615 | / | / | VapB | 抗毒素 |
| BAA17391 | 893261 | 893590 | 109 | + | 未知蛋白 slr1209 | PF04255 | 未知功能蛋白 (DUF433) | VapB | 抗毒素 |
| BAA17392 | 893583 | 893972 | 129 | + | 未知蛋白 slr1210 | PF18480 | 未知功能结构域 (DUF5615) | VapC | 毒素 |
| BAA17455 | 968487 | 968960 | 157 | - | 未知蛋白 sll1715 | PF01850 | PIN结构域结构域 | VapC | 毒素 |
| BAA17456 | 968893 | 969294 | 133 | - | 未知蛋白 sll1714 | / | / | VapB | 抗毒素 |
| BAA17528 | 1048406 | 1048753 | 115 | - | 未知蛋白 sll1130 | PF02452 | PemK-样毒素 | MazF | 毒素 |
| BAA17529 | 1048750 | 1049016 | 88 | - | 未知蛋白 ssl2245 | / | / | MazE | 抗毒素 |
| BAA17539 | 1056714 | 1056944 | 76 | + | HicB | / | / | HicB | 抗毒素 |
| New ORF4 | 1056487 | 1056717 | 76 | + | New ORF | / | / | HicA | 毒素 |
| BAA17886 | 1444115 | 1445392 | 425 | + | 未知蛋白 slr1998 | PF01494 | FAD 结合结构域 | 未知 | 未知 |
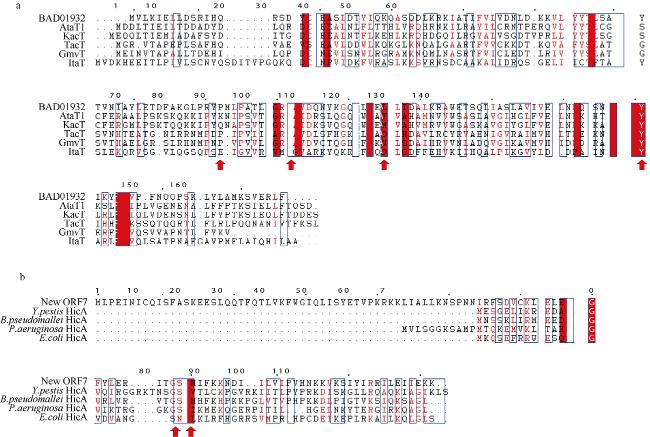
| BAA17887 | 1445425 | 1445835 | 136 | + | 未知蛋白 Gura_0600 | PF07927 | HicA | N-HicA,C-HicB | 抗毒素 |
| BAA18037 | 1617376 | 1617813 | 145 | - | 未知蛋白 sll1411 | PF01934 | 未知功能蛋白 DUF86 | HEPN | 毒素 |
| BAA18038 | 1617806 | 1618096 | 96 | - | 未知蛋白 ssl2749 | PF01909 | 核苷酸转移酶结构域结构域 | MNT | 抗毒素 |
| BAA18040 | 1620037 | 1620222 | 61 | + | 未知蛋白 ssr3570 | / | / | YoeB | 毒素 |
| BAA18041 | 1620276 | 1620539 | 87 | + | prevent-host-death family protein | PF02604 | Phd_YefM抗毒素 | YefM | 抗毒素 |
| 起始位点 | 终止位点 | 蛋白长度(氨基酸) | 正负链 | 基因功能描述 | 蛋白家族编号 | 蛋白家族描述 | xtalpred 软件分析结果 | 毒素/抗毒素 | |
| BAA18080 | 1660069 | 1660287 | 72 | + | 未知蛋白 ssr2201 | / | / | VapB | 抗毒素 |
| BAA18081 | 1660277 | 1660711 | 144 | + | 未知蛋白 slr1327 | PF13470 | PIN结构域 | VapC | 毒素 |
| BAA18337 | 1933468 | 1933854 | 128 | - | 未知蛋白 SYNGTS_1762 | PF01850 | PIN结构域 | VapC | 毒素 |
| BAA18338 | 1933854 | 1934060 | 68 | - | 未知蛋白 SYNGTS_1763 | PF10047 | 未知功能蛋白 (DUF2281) | VapB | 抗毒素 |
| New ORF5 | 2084922 | 2085206 | 94 | + | New ORF | / | / | BrnT | 毒素 |
| New ORF6 | 2085136 | 2085417 | 93 | + | New ORF | PF07927 | HicA | BrnA | 抗毒素 |
| BAA18559 | 2193591 | 2193812 | 73 | - | 未知蛋白 MAE_17890 | PF15919 | HicB样抗毒素 | HicB | 抗毒素 |
| New ORF7 | 2193813 | 2194157 | 114 | - | New ORF | PF07927 | HicA, | HicA | 毒素 |
| New ORF8 | 3341880 | 3342128 | 82 | - | New ORF | / | / | HicB | 抗毒素 |
| BAA18682 | 3342125 | 3342442 | 105 | - | 未知蛋白 MAE_13550 | PF13470 | PIN结构域 | VapC | 毒素 |
| BAA18687 | 3347941 | 3348201 | 86 | + | 未知蛋白 SYNGTS_2983 | / | / | VapB | 抗毒素 |
| BAA18688 | 3348191 | 3348553 | 120 | + | 未知蛋白 SYNGTS_2984 | PF06296 | RelE / RelB系统毒素 | RelE | 毒素 |
| BAA18757 | 3428962 | 3429171 | 69 | + | 未知蛋白 MAE_40070 | PF15919 | HicB样抗毒素 | HicB | 抗毒素 |
| New ORF9 | 3429171 | 3429398 | 75 | + | New ORF | / | / | HicA | 毒素 |
| BAB61866 | 2074694 | 2074957 | 87 | + | prevent-host-death family protein (plasmid) | PF02604 | Phd_YefM类抗毒素 | YefM 抗毒素 | 抗毒素 |
| BAB61867 | 2085826 | 2086110 | 94 | + | prevent-host-death family protein | PF02604 | Phd_YefM类抗毒素 | YefM 抗毒素 | 抗毒素 |
| BAD01773 | 1136 | 1540 | 134 | - | 未知蛋白 sll5003 (plasmid) | PF18480 | 未知功能结构域 (DUF5615) | VapC | 毒素 |
| BAD01774 | 1515 | 1826 | 103 | - | 未知蛋白 sll5004 (plasmid) | PF04255 | 未知功能蛋白 (DUF433) | VapB | 抗毒素 |
| BAD01790 | 24181 | 24462 | 93 | + | 未知蛋白 ssr5020 (plasmid) | PF05015 | RelE 样毒素 | HigB-like_毒素 | 毒素 |
| BAD01791 | 24473 | 24787 | 104 | + | 未知蛋白 slr5021 | PF01381 | 螺旋-转角-螺旋 | HigA_抗毒素 | 抗毒素 |
| BAD01835 | 67163 | 67378 | 71 | - | 未知蛋白 ssl5065 | / | / | ParD_抗毒素 | 抗毒素 |
| BAD01836 | 67362 | 67976 | 204 | - | ParA 家族 | PF01656 | CobQ/CobB/MinD/ParA 核苷酸结合结构域 | ParA_毒素 | 毒素 |
| BAD01864 | 92805 | 93236 | 143 | - | 质粒稳定蛋白 | PF01850 | PIN结构域 | VapC | 毒素 |
| BAD01865 | 93233 | 93520 | 95 | - | 未知蛋白 ssl5095 | PF02604 | Phd_YefM类抗毒素 | VapB | 抗毒素 |
| BAD01871 | 96124 | 96561 | 145 | + | 未知蛋白 Ajs_1570 | / | / | 未知 | 未知 |
| BAD01872 | 96554 | 97156 | 200 | + | 未知蛋白 BURPS1106A_2452 | PF08843 | 核苷酸转移酶 AbiEii毒素 | AbiEii 毒素 | 毒素 |
| BAD01886 | 107105 | 107527 | 140 | + | 未知蛋白 slr5116 (plasmid) | PF13470 | PIN结构域 | VapC | 毒素 |
| BAD01887 | 107531 | 107746 | 71 | + | 未知蛋白 ssr5117 | PF05534 | HicB 家族 | VapB | 抗毒素 |
| BAD01905 | 1399 | 1827 | 142 | - | 质粒稳定蛋白 | PF01850 | PIN结构域 | VapC | 毒素 |
| BAD01906 | 1799 | 2032 | 77 | - | 未知蛋白 MYO_450 | / | / | VapB | 抗毒素 |
| BAD01908 | 3041 | 3364 | 107 | - | 未知蛋白 sll7006 | PF05534 | HicB 家族 | HicB | 抗毒素 |
| BAD01909 | 3367 | 3591 | 74 | - | 未知蛋白 ssl7007 | / | / | VapC | 毒素 |
| BAD01932 | 28850 | 29353 | 167 | - | 未知蛋白 sll7030 | PF13508 | 乙酰基转移酶(GNAT)结构域 | GNAT 毒素 | 毒素 |
| BAD01933 | 29337 | 29639 | 100 | - | 未知蛋白 sll7031 | PF08681 | 未知功能蛋白 (DUF1778) | GNAT 抗毒素 | 抗毒素 |
| BAD01940 | 35891 | 36178 | 95 | - | 未知蛋白 ssl7038 | / | / | HigA 抗毒素 | 抗毒素 |
| BAD01941 | 36189 | 36479 | 96 | - | 未知蛋白 ssl7039 | PF05973 | 噬菌体衍生蛋白 Gp49 样(DUF891) | HigB 毒素 | 毒素 |
| BAD01942 | 36597 | 36848 | 83 | + | 未知蛋白 ssr7040 | PF04014 | MazE抗毒素 | MazE | 抗毒素 |
| BAD01943 | 36839 | 37201 | 120 | + | 未知蛋白 slr7041 | PF02452 | PemK-样, MazF-样毒素 | MazF | 毒素 |
| BAD01947 | 40013 | 40234 | 73 | - | 未知蛋白 ssl7045 | / | / | 未知 | 未知 |
| 起始位点 | 终止位点 | 蛋白长度(氨基酸) | 正负链 | 基因功能描述 | 蛋白家族编号 | 蛋白家族描述 | xtalpred 软件分析结果 | 毒素/抗毒素 | |
| BAD01948 | 40245 | 40514 | 89 | - | 未知蛋白 sll8027 | PF13470 | PIN结构域 | VapC | 毒素 |
| BAD01997 | 93673 | 94188 | 171 | + | 未知蛋白 slr7095 | PF13671 | AAA结构域 | 未知 | 毒素 |
| BAD01998 | 94198 | 94611 | 137 | + | 未知蛋白 slr7096 | / | / | 未知 | 未知 |
| BAD02012 | 3836 | 4393 | 185 | - | 未知蛋白 sll8004 | PF13671 | AAA结构域 | Zeta_毒素 | 毒素 |
| BAD02013 | 4365 | 4655 | 96 | - | 未知蛋白 ssl2749 | PF01909 | 核苷酸转移酶结构域 | MNT | 抗毒素 |
| BAD02015 | 6043 | 6393 | 116 | - | 未知蛋白 sll8007 | PF18478 | PIN 样结构域 | VapC | 毒素 |
| BAD02016 | 6377 | 6601 | 74 | - | 未知蛋白 ssl8008 | PF04255 | 未知功能蛋白 (DUF433) | VapB | 抗毒素 |
| BAD02019 | 8821 | 9195 | 124 | - | 未知蛋白 sll8011 | PF18480 | 未知功能结构域 (DUF5615) | VapC | 毒素 |
| BAD02020 | 9205 | 9621 | 138 | - | 未知蛋白 sll8012 | / | / | VapB | 抗毒素 |
| BAD02021 | 9713 | 9940 | 75 | + | 未知蛋白 ssr8013 | PF10047 | 未知功能蛋白 (DUF2281) | VapB | 抗毒素 |
| BAD02022 | 9948 | 10373 | 141 | + | 未知蛋白 slr8014 | PF13470 | PIN结构域 | VapC | 毒素 |
| BAD02023 | 10395 | 10994 | 199 | + | ParA 家族 | PF01656 | CobQ/CobB/MinD/ParA 核苷酸结合结构域 | ParA | 毒素 |
| BAD02024 | 10998 | 11867 | 289 | + | 质粒分配蛋白 | PF02195 | ParB-样核酸酶结构域 | ParB | 抗毒素 |
| BAD02035 | 23434 | 23859 | 141 | - | 未知蛋白 sll8027 | PF13470 | PIN结构域 | VapC | 毒素 |
| BAD02036 | 23856 | 24077 | 73 | - | 未知蛋白 ssl8028 | / | / | VapB | 抗毒素 |
| BAD02106 | 47396 | 47815 | 139 | + | PilT | PF01850 | PIN结构域 | VapC | 毒素 |
| BAD02107 | 47827 | 51129 | 1100 | + | I型限制修饰系统M亚基 | PF07669 | Eco57I限制性修饰甲基化酶 | VapB | 抗毒素 |
| BAD02149 | 87011 | 87274 | 87 | - | 未知蛋白 ssl6092 | / | / | VapC | 抗毒素 |
| BAD02150 | 87258 | 87854 | 198 | - | ParA 家族 | PF01656 | CobQ/CobB/MinD/ParA 核苷酸结合结构域 | VapB | 抗毒素 |
| BAD02157 | 93820 | 94140 | 106 | + | 未知蛋白 slr6100 | PF09907 | HigB_毒素, RelE-样毒素 | HigB_毒素 | 毒素 |
| BAD02158 | 94281 | 94700 | 139 | + | 未知蛋白 slr6101 | / | / | HigA_抗毒素 | 抗毒素 |
附表3 聚球藻 WH7803 毒素-抗毒素系统预测信息Tab. S3 Prediction information of toxin-antitoxin systems in Synechococcus sp. WH7803 |
| 基因编号(ensembl数据库) | 起始位点 | 终止位点 | 蛋白长度(氨基酸) | 正负链 | 基因功能描述 | 蛋白家族编号 | 蛋白家族描述 | xtalpred 软件分析结果 | 毒素/抗毒素 | |
|---|---|---|---|---|---|---|---|---|---|---|
| SynWH7803_0100 | 102513 | 102677 | 54 | + | NTP_transf_2 结构域蛋白 | / | 未知 | 未知 | 毒素 | |
| SynWH7803_0101 | 102623 | 102820 | 65 | + | RelA 和 Spot 样 ppGpp 合成酶和 水解酶 | / | NT_Pol-β-样 超家族 | MNT | 抗毒素 | |
| SynWH7803_0976 | 902407 | 902246 | 53 | + | 未表征的膜蛋白 | / | PIN | 未知 | 抗毒素 | |
| SynWH7803_0977 | 902875 | 902492 | 127 | + | 保守的假定蛋白保守的假定蛋白 | PF11964 | SpoIIAA-样 | SpoIIAA-样蛋白 | 未知 | |
| SynWH7803_1086 | 996599 | 995259 | 446 | - | 保守的假定蛋白保守的假定蛋白 | PF14403 | Circularly permuted ATP-grasp type 2 | 谷胱甘肽精氨酸合酶 | 未知 | |
| SynWH7803_1087 | 997032 | 996586 | 148 | - | 保守的假定蛋白保守的假定蛋白 | / | / | 未知 | 抗毒素 | |
| SynWH7803_1088 | 997508 | 997032 | 158 | - | 分泌的五肽重复蛋白 | PF00805 | 五肽重复序列 (8 份) | 未知 | 未知 | |
| SynWH7803_1121 | 1027292 | 1026795 | 165 | - | 预测的 rRNA 甲基化酶 | PF00588 | SpoU rRNA 甲基化酶家族 | RNA甲基化酶 | 毒素 | |
| SynWH7803_1122 | 1027843 | 1027289 | 184 | - | 双功能腺苷钴胺生物合成蛋白 | PF02283 | 甲酰胺激酶 | 腺苷钴酰胺激酶/腺苷钴酰胺磷酸鸟苷转移酶 | 未知 | |
| SynWH7803_1603 | 1475142 | 1474984 | 52 | + | 假定蛋白 | / | / | ATP依赖性RNA解旋酶 | 未知 | |
| SynWH7803_1604 | 1475671 | 1475198 | 157 | + | 未表征的保守 分泌蛋白 | / | Gp49 | 未知 | 毒素 | |
| SynWH7803_1605 | 1476140 | 1475679 | 153 | + | SufE 样蛋白, 可能参与 Fe-S 中心组装 | PF02657 | Fe-S代谢相关域 | Cpta_toxin | 毒素 | |
| 基因编号(ensembl数据库) | 起始位点 | 终止位点 | 蛋白长度(氨基酸) | 正负链 | 基因功能描述 | 蛋白家族编号 | 蛋白家族描述 | xtalpred 软件分析结果 | 毒素/抗毒素 | |
| SynWH7803_1606 | 1477492 | 1476176 | 438 | + | 高丝氨酸脱氢酶 | PF03447 | 高丝氨酸脱氢酶, NAD 结合结构域 | 高丝氨酸脱氢酶 | 未知 | |
| SynWH7803_1700 | 1554939 | 1554607 | 110 | - | 假定蛋白 | PF13469 | 磺基转移酶家族 | 磺基转移酶家族 | 未知 | |
| SynWH7803_1701 | 1555576 | 1554899 | 225 | - | 假定蛋白 | PF13469 | 磺基转移酶家族 | 磺基转移酶家族 | 毒素 | |
| SynWH7803_1762 | 1616798 | 1615596 | 400 | + | 整合膜蛋白(PIN 域超家族) | / | PIN_5 | VapC | 毒素 | |
| SynWH7803_1763 | 1617500 | 1616829 | 223 | + | ATP 依赖性 Clp 蛋白酶的蛋白酶亚基 | PF00574 | Clp 蛋白酶 | Clp 蛋白酶 | 未知 | |
| SynWH7803_1944 | 1778918 | 1779145 | 75 | + | 假定蛋白SynWH7803_1944 | PF02604 | Phd YefM抗毒素 | Phd YefM_超家族抗毒素 | 抗毒素 | |
| SynWH7803_1945 | 1779145 | 1779555 | 136 | + | 假定蛋白 SynWH7803_1945 | PF01850 | PIN 结构域 | VapC | 毒素 | |
| SynWH7803_2509 | 2341171 | 2340317 | 284 | - | 甲酰基四氢叶酸去甲酰基酶 | PF00551 | 甲酰基转移酶 | 甲酰基转移酶 | 未知 | |
| SynWH7803_2510 | 2341482 | 2341222 | 86 | - | 预测的转录调节因子 | PF02195 | ParB 样核酸酶 结构域 | ParBc | 抗毒素 | |
表1 本文选取的7对毒素-抗毒素系统预测信息。Tab. 1 Prediction information of seven TA systems selected in this paper |
| 菌株名称 | 基因编号 | 起始位置 | 终止位置 | 蛋白长度/aa | TA信息 | TA 类型预测 |
|---|---|---|---|---|---|---|
| 集胞藻PCC6803 | BAD01871 | 96124 | 96561 | 145 | x | |
| BAD01872 | 96554 | 97156 | 200 | T | AbiEii家族毒素 | |
| BAD01932 | 28850 | 29353 | 167 | x | GNAT家族毒素 | |
| BAD01933 | 29337 | 29639 | 100 | A | GNAT家族抗毒素 | |
| BAD01997 | 93673 | 94188 | 171 | T | NTP水解酶家族毒素 | |
| BAD01998 | 94198 | 94611 | 137 | x | ||
| BAA17887 | 1445425 | 1445835 | 136 | A | HicA和HicB融合蛋白 | |
| New ORF 7 | 2193818 | 2194157 | 114 | T | HicAB家族毒素 | |
| BAA18559 | 2193591 | 2193812 | 73 | A | HicAB家族抗毒素 | |
| 聚球藻WH7803 | SynWH7803_1121 | 1026795 | 1027292 | 165 | T | 甲基转移酶 |
| SynWH7803_1122 | 1027289 | 1027843 | 184 | x | ||
| SynWH7803_1700 | 1554607 | 1554939 | 110 | x | 硫转移酶家族 | |
| SynWH7803_1701 | 1554899 | 1555576 | 225 | T | 硫转移酶家族 |
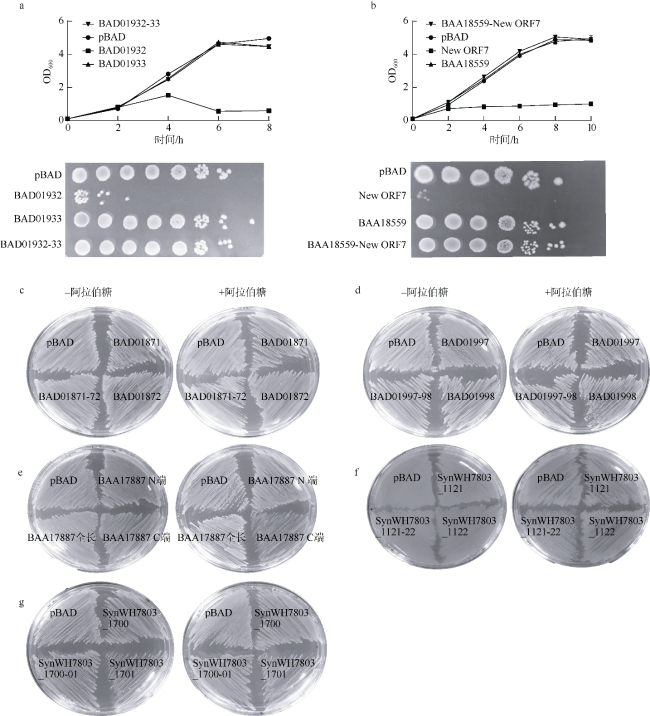
图2 在大肠杆菌BW25113 中鉴定所选7对毒素-抗毒素系统的毒性a、b分别为BAD01932-33和BAA18559-New ORF7毒素-抗毒素系统的生长曲线(OD600)和添加诱导剂4h后各重组菌株的菌落形成单位; c—g分别为BAD01871-72、BAD01997-98、BAA17887、SynWH7803_1121-22 和 SynWH7803_1700-01的毒性验证结果 Fig. 2 Identification of seven candidate TA systems in E. coli BW25113. a-b: The growth curve and colony-forming unit of BAD01932-33 (a) and BAA18559-New ORF7 (b) TA systems. c-g: Toxicity test of BAD01871-72 (c), BAD01997-98 (d), BAA1788 (e), SynWH7803_1121-22 (f), and SynWH7803_1700-01 (g) |
图5 采用大肠杆菌BL21(DE3)表达和纯化GNAT家族抗毒素蛋白BAD01933 (a)、毒素BAD01932与抗毒素合体(b)、HicAB家族抗毒素蛋白BAA18559(c)和毒素New ORF7与抗毒素合体(d)M: 蛋白marker; 1: 未诱导全菌液; 2: 诱导后全菌液; 3: 细菌超声破碎后离心所得到上清液; 4: 包涵体; 5: 上清液流穿Ni柱未结合的部分; 6-7: 蛋白洗脱管 1 和2 号 Fig. 5 Expression and purification of BAD01932-BAD01933 (a, b) and BAA18559- New ORF7 antitoxin and TA complex (c, d). M: Protein marker; 1: whole cell lysate of uninduced cells; 2: whole cell lysate of induced cells; 3: the supernatant of ultrasonication after centrifugation; 4: the inclusion body after ultrasonication and centrifugation; 5: the unbound part from Ni-NTA column; 6-7: the eluted fractions from the Ni-NTA column |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}