
基于形态学和DNA分子鉴定的珠江口浮游动物群落结构比较研究
|
尹天齐(1996—), 女, 河南省平顶山市人, 硕士研究生, 从事浮游动物生态学研究。email: |
Copy editor: 林强
收稿日期: 2021-11-05
修回日期: 2021-12-15
网络出版日期: 2021-12-23
基金资助
广东省自然科学基金面上项目(2022A1515011387)
广东省自然科学基金面上项目(2021A1515010814)
Comparative study on zooplankton community structure in Pearl River Estuary based on morphological and DNA identification
Copy editor: LIN Qiang
Received date: 2021-11-05
Revised date: 2021-12-15
Online published: 2021-12-23
Supported by
General project of Guangdong Natural Science Foundation(2022A1515011387)
General project of Guangdong Natural Science Foundation(2021A1515010814)
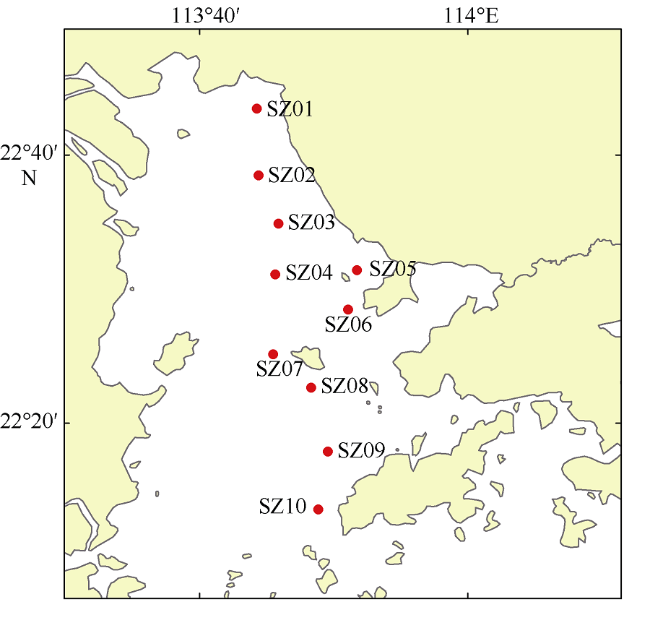
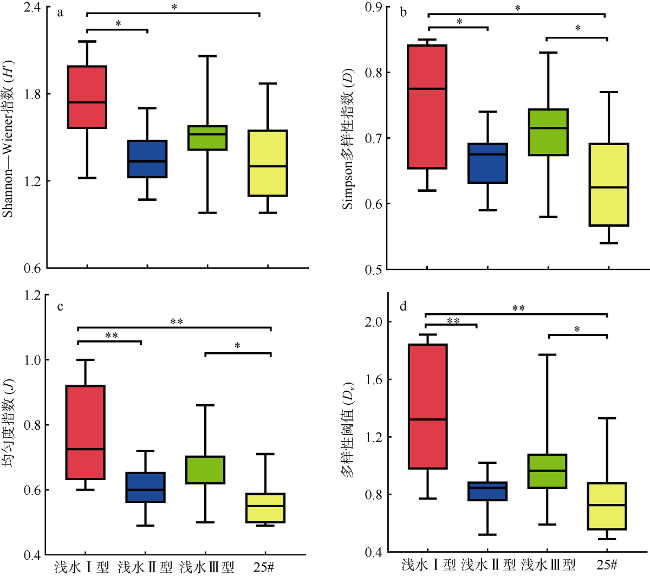
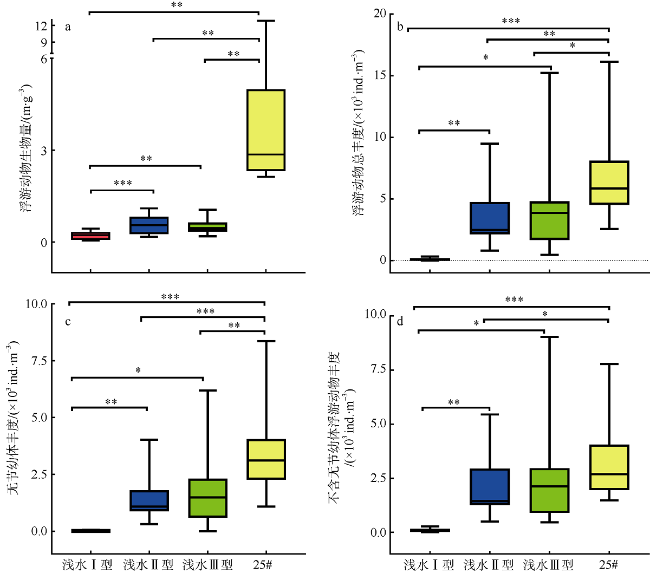
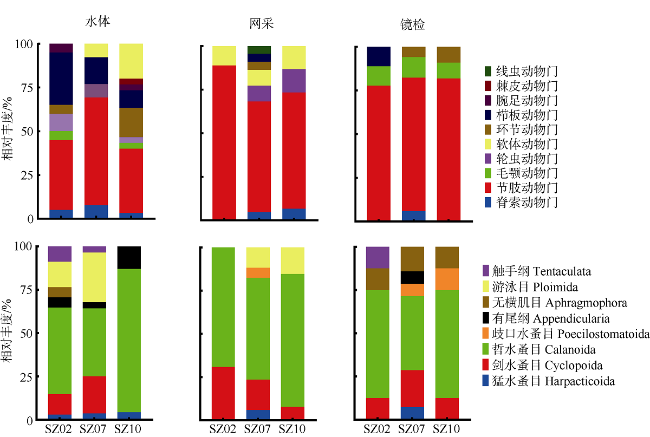
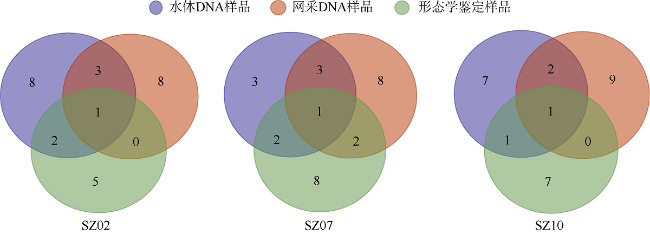
于2019年12月使用4种不同型号网具采集了珠江口浮游动物, 进行形态学鉴定和DNA分子鉴定, 分析珠江口浮游动物的群落结构特征, 并比较不同调查方法对浮游动物丰度和生物量结果的影响。形态学镜检鉴定浮游动物36种(类), 其中浮游幼虫6类。浅水I型浮游生物网采集的浮游动物平均丰度为115±96ind.·m-3, 平均生物量为0.21±0.14g·m-3; 浅水Ⅱ型浮游生物网采集的浮游动物平均丰度为3536±2444ind.·m-3, 平均生物量为0.56±0.33g·m-3; 浅水Ⅲ型浮游生物网采集的浮游动物平均丰度为4314±4172ind.·m-3, 平均生物量为0.50±0.25g·m-3; 25#浮游生物网采集的浮游动物平均丰度为6741±3826ind.·m-3, 平均生物量为4.33±3.42g·m-3。研究结果表明网具孔径大小对浮游动物研究结果具有重要影响, 三个站点水体DNA样品注释出15种浮游动物; 使用浅水Ⅱ型网采集的DNA样品注释出19种浮游动物; 镜检样品鉴定浮游动物17种。水体DNA样品能检测出更多的微型浮游动物如原生动物等; 网采样品能过滤更多的水样, 有利于采集更多的大中型浮游动物, 更能充分反映优势类群如桡足类的种类和数量。研究结果表明, 水体DNA可检出浮游幼虫和原生动物等较难镜检鉴别的种类, 采用不同型号网具采集浮游动物可以更全面地反映研究海域浮游动物的群落结构特征。多种调查方法的结合有助于全面了解研究海域的生态环境状况。
尹天齐 , 王庆 , 杨宇峰 , 岑竞仪 . 基于形态学和DNA分子鉴定的珠江口浮游动物群落结构比较研究[J]. 热带海洋学报, 2022 , 41(3) : 172 -185 . DOI: 10.11978/2021151
Zooplankton samples in the Pearl River Estuary were collected in December 2019 using four different types of plankton nets. We analyzed the characteristics of zooplankton community structure and compared the effects of different investigation methods on the study of zooplankton community structure based on morphological identification and DNA monitoring. Thirty-six species of zooplankton including six species of planktonic larvae were identified by morphological identification. The average abundance of zooplankton collected by the I type of plankton net for the shallow water was 115±96 ind·m-3, and the average biomass was 0.21±0.14g·m-3. The average abundance of zooplankton collected by the Ⅱ type of plankton net for the shallow water was 3536±2444ind·m-3, and the average biomass was 0.56±0.33 g·m-3. The average abundance of zooplankton collected by the Ⅲ type of plankton net for the shallow water was 4314±4172 ind.·m-3, and the average biomass was 0.50±0.25 g·m-3. The average abundance of zooplankton collected by 25# plankton net was 6741±3826 ind·m-3, and the average biomass was 4.33±3.42 mg·m-3. Results show that the mesh size of plankton net has an important impact on the results of zooplankton. Water DNA samples were collected from three sites annotated 15 species of zooplankton; DNA samples were collected by the Ⅱ type of plankton net from three sites annotated 19 species of zooplankton; and 17 species of zooplankton were identified by morphology at the three sites. Results show that water DNA samples can detect micro-zooplankton, such as protozoa. DNA samples collected by the plankton net can filter much water, which is conducive to collecting more macro- and meso-sized zooplankton, and can fully reflect the species and abundance of dominant groups such as copepods. Water DNA samples can be used to detect species that are difficult to be identified, such as planktonic larvae and protozoa. Zooplankton collected with different types of plankton nets can comprehensively reflect the characteristics of zooplankton community structure in the study area. The combination of various investigation methods is helpful to comprehensively understand the ecological environment in the study area.
Key words: Pearl River Estuary; zooplankton; particle size; DNA identification
表1 珠江口浮游动物的种类组成Tab. 1 Species composition of zooplankton in the Pearl River Estuary |
| 类别 | 种类 | Ⅰ型网 | Ⅱ型网 | Ⅲ型网 | 25#网 |
|---|---|---|---|---|---|
| 桡足类 | 红纺锤水蚤(Acartia erythraea) | + | + | + | |
| 太平洋纺锤水蚤(Acartia pacifica) | + | + | + | + | |
| 刺尾纺锤水蚤(Acartia spinicauda) | + | + | + | + | |
| 中华异水蚤(Acartiella sinensis) | + | + | + | + | |
| 拟矮隆水蚤(Bestiolina similis) | + | + | |||
| 叉胸刺水蚤(Centropages furcatus) | + | ||||
| 微刺哲水蚤(Canthocalanus pauper) | + | + | |||
| 精致真刺水蚤(Euchaeta concinna) | + | + | |||
| 卵型光水蚤(Lucicutia ovalis) | + | ||||
| 强额孔雀哲水蚤(Parvocalanus crassirostris) | + | + | + | + | |
| 小拟哲水蚤(Paracalanus parvus) | + | ||||
| 亚强次真哲水蚤(Subeucalanus subcrassus) | + | + | + | + | |
| 异尾宽水蚤(Temora discaudata) | + | ||||
| 锥形宽水蚤(Temora turbinata) | + | + | + | + | |
| 瘦歪水蚤(Tortanus gracilis) | + | ||||
| 近缘大眼剑水蚤(Corycaeus affinis) | + | ||||
| 平大眼水蚤(Corycaeus dahli) | + | + | |||
| 近邻剑水蚤(Cyclops vicinus) | + | + | + | ||
| 短角长腹剑水蚤(Oithona brevicornis) | + | + | + | ||
| 小长腹剑水蚤(Oithona nana) | + | + | + | + | |
| 瘦长毛猛水蚤(Macrosetella gracilis) | + | + | + | ||
| 小毛猛水蚤(Microsetella norvegica) | + | + | + | + | |
| 水母类 | 半口壮丽水母(Aglaura hemistoma) | + | + | + | |
| 不列颠高手水母(Bougainvillia britannica) | + | + | + | ||
| 双生水母(Diphyes chamissonis) | + | + | + | ||
| 短柄和平水母(Eirene brevistylus) | + | + | + | ||
| 球型侧腕水母(Pleurobrachia globosa) | + | + | + | ||
| 毛颚类 | 百陶带箭虫(Zonosagitta bedoti) | + | + | + | + |
| 肥胖软箭虫(Flaccisagitta enflata) | + | + | + | + | |
| 被囊类 | 异体住囊虫(Oikopleura longicaudata) | + | + | + | |
| 浮游幼虫 | 磷虾幼虫(Euphausiacea larva) | + | + | + | + |
| 蔓足类幼虫(Cirripedia larva) | + | + | + | + | |
| 莹虾幼虫(Lucifer larva) | + | + | + | + | |
| 长尾类幼虫(Macruran larva) | + | + | + | ||
| 多毛类幼虫(Polychaeta larva) | + | + | + | + | |
| 鱼卵 | 鱼卵(fish eggs) | + | + | + |
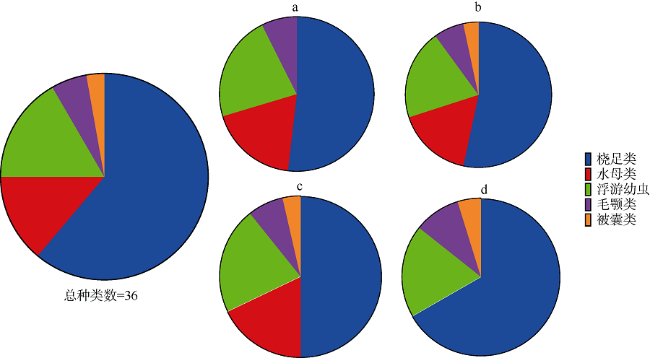
图2 珠江口浮游动物类群的物种丰富度a. 浅水Ⅰ型网(共27种); b. 浅水Ⅱ型网(共30种); c. 浅水Ⅲ型网(共28种); d. 25#浮游生物网(共21种) Fig. 2 Species richness of zooplankton groups in the Pearl River Estuary. a. Type I net for shallow water (27 species); b. Type Ⅱ net for shallow water (30 species); c. Type Ⅲ net for shallow water (28 species); and d. 25# Plankton net (21 species) |
表2 不同网具采集的浮游动物种类优势度Tab. 2 Species dominance of zooplankton collected using different types of nets |
| 种类 | 浅水Ⅰ型网 | 浅水Ⅱ型网 | 浅水Ⅲ型网 | 25#浮游生物网 | ||||
|---|---|---|---|---|---|---|---|---|
| 优势度Y | 平均丰度/(ind·m-3) | 优势度Y | 平均丰度/(ind·m-3) | 优势度Y | 平均丰度/(ind·m-3) | 优势度Y | 平均丰度/(ind·m-3) | |
| 强额孔雀哲水蚤(Parvocalanus crassirostris) | 0.04 | 18 | 0.34 | 1070 | 0.15 | 666 | 0.24 | 2182 |
| 太平洋纺锤水蚤(Acartia pacifica) | 0.10 | 46 | <0.02 | / | <0.02 | / | <0.02 | / |
| 短角长腹剑水蚤(Oithona brevicornis) | <0.02 | / | <0.02 | / | 0.05 | 193 | 0.43 | 640 |
| 微刺哲水蚤(Canthocalanus pauper) | <0.02 | / | <0.02 | / | 0.04 | 1007 | <0.02 | / |
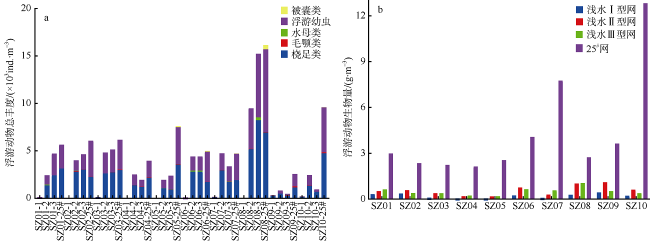
图4 珠江口浮游动物丰度和生物量的空间变化a. 浮游动物丰度; b. 浮游动物生物量。站点后缀-1、-2、-3和-25#分别表示在该站点使用浅水Ⅰ型网、Ⅱ型网、Ⅲ型网和25#浮游生物网采集的浮游动物样品 Fig. 4 Spatial variation of zooplankton abundance and biomass in the Pearl River Estuary (a) zooplankton abundance; and (b) zooplankton biomass. Site suffixes -1, -2, -3, and -25#, respectively, represent zooplankton samples collected by shallow water type I net, type Ⅱ net, type III net, and 25# plankton net at the sampling site |
表3 DNA样品序列数和OTU数Tab. 3 The number of sequence and OTU in the DNA samples |
| 站位 | 原始序列数 | 高质量序列数 | OTU数 | |||
|---|---|---|---|---|---|---|
| 水体 | 网采 | 水体 | 网采 | 水体 | 网采 | |
| SZ02 | 43730 | 71598 | 43130 | 63055 | 1112 | 75 |
| SZ07 | 42697 | 64065 | 42450 | 56946 | 494 | 175 |
| SZ10 | 34802 | 59620 | 34671 | 69004 | 817 | 65 |
表4 水体和网采DNA样品测序与形态学鉴定种类Tab. 4 Zooplankton species in the water and net-collected samples through DNA sequencing and morphological identification |
| 种类 | 水体DNA样品OTU数 | 网采DNA样品OTU数 | 镜检种类数 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| SZ02 | SZ07 | SZ10 | SZ02 | SZ07 | SZ10 | SZ02 | SZ07 | SZ10 | |
| 长纺锤水蚤(Acartia longiremis) | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 太平洋纺锤水蚤(Acartia pacifica) | 3 | 3 | 4 | 0 | 0 | 0 | 1 | 1 | 1 |
| 刺尾纺锤水蚤(Acartia spinicauda) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| 中华异水蚤(Acartiella sinensis) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 |
| 拟矮隆水蚤(Bestiolina similis) | 2 | 2 | 2 | 1 | 1 | 1 | 0 | 0 | 0 |
| 叉胸刺水蚤(Centropages furcatus) | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| 胸刺水蚤(Centropages hamatus) | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| 典型胸刺水蚤(Centropages typicus) | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 强额孔雀哲水蚤(Parvocalanus crassirostris) | 5 | 1 | 6 | 2 | 2 | 2 | 1 | 1 | 1 |
| 伪镖水蚤(Pseudodiaptomus euryhalinus) | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 |
| 指状伪镖水蚤(Pseudodiaptomus inopinus) | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 中华华哲水蚤(Sinocalanus sinensis) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 强次真哲水蚤(Subeucalanus crassus) | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 |
| 亚强次真哲水蚤(Subeucalanus subcrassus) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 |
| 异尾宽水蚤(Temora discaudata) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| 锥形宽水蚤(Temora turbinata) | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 |
| 瘦歪水蚤(Tortanus gracilis) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| 近缘大眼剑水蚤(Corycaeus affinis) | 0 | 0 | 0 | 2 | 2 | 1 | 0 | 1 | 0 |
| 平大眼水蚤(Corycaeus dahli) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| 近邻剑水蚤(Cyclops vicinus) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| 半剑水蚤(Hemicyclops sp.) | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 半剑水蚤(Hemicyclops tanakai) | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 真刺唇角水蚤(Labidocera euchaeta) | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 |
| 短角长腹剑水蚤(Oithona brevicornis) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| 戴维斯长腹剑水蚤(Oithona davisae) | 2 | 2 | 2 | 1 | 1 | 0 | 0 | 0 | 0 |
| 小长腹剑水蚤(Oithona nana) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| 长腹剑水蚤(Oithona sp.) | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 0 |
| 矮小拟镖剑水蚤(Paracyclopina nana) | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 |
| 小毛猛水蚤(Microsetella norvegica) | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 0 |
| 拟双倍猛水蚤(Paramphiascella fulvofasciata) | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 三角大吉猛水蚤(Tachidius triangularis) | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 箭虫(无横肌目)(Aphragmophora) | 2 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 百陶带箭虫(Zonosagitta bedoti) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| 肥胖软箭虫(Flaccisagitta enflata) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| 卵形无柄轮虫(Ascomorpha ovalis) | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 |
| 萼花臂尾轮虫(Brachionus calyciflorus) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 梳状疣毛轮虫(Synchaeta pectinata) | 1 | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 颤动毛轮虫(Synchaeta tremula) | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 |
| 球型侧腕水母(Pleurobrachia globosa) | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| 斑点蝶水母(Ocyropsis maculata) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 住囊虫(Oikopleura sp.) | 2 | 2 | 3 | 0 | 0 | 0 | 0 | 1 | 0 |
注: 水体DNA样品和网采DNA样品中的数字表示归类到种属的浮游动物OTU条数, 未列入不能归类到种属的浮游动物OTU |
表5 珠江口浮游动物丰度和生物量调查结果的比较Tab. 5 Comparison of zooplankton abundance and biomass in the Pearl River Estuary among different investigations |
| 采样时间 | 网具 | 丰度/(ind.·m-3) | 生物量/(mg·m-3) | 珠江口 |
|---|---|---|---|---|
| 1959 | 浅水I型网 | / | 58~101 | 全国海洋普查(张达娟 等, 2008) |
| 1960 | 浅水I型网 | / | 33 | 全国海洋普查(张达娟 等, 2008) |
| 1981 | 浅水I型网 | / | 213~239 | 中国海湾志编纂委员会, 1998 |
| 1995 | 浅水I型网 | / | 2423 | 郑奕麟, 1995 |
| 1996 | 浅水I型网 | 5.01~606.5 | / | 刘玉 等, 2001 |
| 2002—2003 | 浅水I型网 | 700~1131 | 203~382 | 李开枝 等, 2005 |
| 2002—2003 | 浅水I型网 | 73~185 | / | 李开枝 等, 2007 |
| 2004 | 浅水I型网 | / | 152.34~571.54 | 张达娟 等, 2008 |
| 2005 | 浅水I型网 | / | 444.77~656.18 | 张达娟 等, 2008 |
| 2006 | 浅水I型网 | / | 712.94~874.5 | 张达娟 等, 2008 |
| 2006—2007 | 浅水I型网 | 2288~28013 | 10.03~132.95 | 高原 等, 2008 |
| 2009 | 浅水I型网 | 423.46 | / | 国家监测中心, 2009 |
| 2012—2013 | 浅水I型网 | / | 20~375 | 彭鹏飞 等, 2015 |
| 2015 | 浅水I型网 | 20.5~1035 | 2.5~417 | 徐姗楠 等, 2017 |
| 2004 | 浅水Ⅱ型网 | 3900~13000 | / | Tan et al, 2004 |
| 2009 | 浅水Ⅱ型网 | 4131.92 | / | 吴玲玲 等, 2012 |
| 2015 | 浅水Ⅱ型网 | 36~896 | / | 刘华雪 等, 2016 |
| 2013—2014 | 浅水Ⅱ型网 | 72.90~35128.57 | / | 黄彬彬 等, 2017 |
| 1985 | 浅水Ⅲ型网 | 31.37~71.68 | 69~204.73 | 宋盛宪, 1991 |
| 2006—2007 | 25#浮游生物网 | 1430~154450 | 6.3~277.3 | 高原 等, 2008 |
| 2019 | 浅水I型网 | 9~331 | 45~444 | 本研究 |
| 2019 | 浅水Ⅱ型网 | 811~9474 | 170~1100 | 本研究 |
| 2019 | 浅水Ⅲ型网 | 460~15221 | 190~1050 | 本研究 |
| 2019 | 25#浮游生物网 | 2560~16140 | 2130~12820 | 本研究 |
| [1] |
陈清潮, 黄良民, 尹建强, 等, 1994. 南沙群岛海区浮游动物多样性研究[M]. 北京: 海洋出版社: 42-50. (in Chinese)
|
| [2] |
国家海洋局南海环境监测中心 2009. 2009年珠江口生态监控区监测报告[R]. 广州: 国家海洋局南海环境监测中心. (in Chinese)
|
| [3] |
高原, 赖子尼, 王超, 等, 2008. 2006年夏季珠江口浮游动物群落结构特征分析[J]. 南方水产, 4(1): 10-15.
|
| [4] |
黄彬彬, 郑淑娴, 蔡伟叙, 等, 2017. 珠江口枯水期和丰水期中小型桡足类种类组成、丰度分布及其与环境因子的关系[J]. 厦门大学学报(自然科学版), 56(6): 852-858.
|
| [5] |
李开枝, 尹健强, 黄良民, 等, 2005. 珠江口浮游动物的群落动态及数量变化[J]. 热带海洋学报, 24(5): 60-68.
|
| [6] |
李开枝, 尹健强, 黄良民, 等, 2007. 珠江口浮游桡足类的生态研究[J]. 生态科学, 26(2): 97-102.
|
| [7] |
李开枝, 柯志新, 王军星, 等. 2021. 西沙群岛珊瑚礁海域浮游动物群落结构初步分析[J]. 热带海洋学报, 41(2): 121-131.
|
| [8] |
连光山, 王彦国, 孙柔鑫, 等, 2018. 中国海洋浮游桡足类多样性[M]. 北京: 海洋出版社: 40-747.
|
| [9] |
连喜平, 谭烨辉, 刘永宏, 等, 2013. 两种浮游生物网对南海北部浮游动物捕获效率的比较[J]. 热带海洋学报, 32(3): 33-39.
|
| [10] |
刘华雪, 许友伟, 陈作志, 等, 2016. 水母旺发对珠江口鱼类资源量的影响[J]. 热带海洋学报, 35(6): 68-73.
|
| [11] |
刘玉, 李适宇, 董燕红, 等, 2001. 珠江口伶仃水道浮游生物及底栖动物群落特征分析[J]. 中山大学学报(自然科学版), 40(S2): 114-118.
|
| [12] |
彭鹏飞, 李绪录, 蔡钰灿, 2015. 珠江口万山群岛海域秋春季浮游动物的分布特征及其与环境因子的关系[J]. 安徽农业科学, 43(18): 170-174.
|
| [13] |
司悦悦, 2018. 珠江口及南海中北部中型浮游动物摄食生态与繁殖研究[D]. 厦门大学.
|
| [14] |
宋盛宪, 1991. 珠江口浮游生物的初步研究[J]. 海洋渔业, 13(1): 24-27. (in Chinese)
|
| [15] |
孙松, 李超伦, 程方平, 等, 2015. 中国近海常见浮游动物图集[M]. 北京: 海洋出版社: 15-79.
|
| [16] |
王荣, 王克, 2003. 两种浮游生物网捕获性能的现场测试[J]. 水产学报, 27(S1): 98-102.
|
| [17] |
吴玲玲, 朱艾嘉, 郭娟, 等, 2012. 珠江口夏季中、小型浮游动物生态特征研究[J]. 海洋通报, 31(6): 689-694.
|
| [18] |
徐姗楠, 龚玉艳, 詹凤娉, 等, 2017. 珠江口海域南沙段浮游动物群落生态特征[J]. 海洋湖沼通报, (6): 82-90.
|
| [19] |
徐旭华, 黄国林, 胡莉娜, 等, 2021. 一种污染桉树组培苗的霉菌形态学及分子生物学鉴定[J]. 现代园艺, 44(9): 24-25, 30. (in Chinese)
|
| [20] |
徐兆礼, 陈亚瞿, 1989. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J]. 生态学杂志, 8(4): 13-15, 19.
|
| [21] |
杨宇峰, 王庆, 陈菊芳, 等, 2006. 河口浮游动物生态学研究进展[J]. 生态学报, 26(2): 576-585.
|
| [22] |
詹金钰, 呼晓庆, 郑玥熙, 等, 2021. 山西翅果油树上三种重要鳞翅目害虫的形态和分子鉴定及生活史观察[J]. 昆虫学报, 64(5): 618-626.
|
| [23] |
张达娟, 闫启仑, 王真良, 2008. 典型河口浮游动物种类数及生物量变化趋势的研究[J]. 海洋与湖沼, 39(5): 536-540.
|
| [24] |
张武昌, 陶振铖, 赵苑, 等, 2019. 中国海浮游桡足类图谱[M]. 2版. 北京: 科学出版社.
|
| [25] |
郑奕麟, 1995. 珠江口(唐家湾)浮游生物调查[J]. 水产科技, (1): 3-5. (in Chinese)
|
| [26] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会 2008a. GB 17378.1-2007 海洋监测规范第7部分:近海污染生态调查和生物监测 [S]. 北京: 中国标准出版社.
General Administration of Quality Supervision Inspection and Quarantine of the People's Republic of China, Standardization Administration of the People's Republic of China, 2008a. GB 17378.1-2007 The specification for marine monitoring-Part 7: Ecological survey for offshore pollution and biological monitoring[S]. Beijing: Standards Press of China. (in Chinese)
|
| [27] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会 2008b. GB/T 12763.1-2007 海洋调查规范第1部分: 总则[S]. 北京: 中国标准出版社.
General Administration of Quality Supervision Inspection and Quarantine of the People's Republic of China, Standardization Administration of the People's Republic of China, 2008b. GB/T 12763.1-2007 Specifications for oceanographic survey-Part 1: general[S]. Beijing: Standards Press of China. (in Chinese)
|
| [28] |
中国海湾志编纂委员会 1998. 中国海湾志第十四分册[M]. 北京: 海洋出版社, 66-75.
Compilation Committee of China Gulf chronicles 1998, China Bay chronicle Volume 14. Beijing: Ocean Press, 66-75. (in Chinese)
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}