
近165年来中沙环礁中北暗沙滨珊瑚生长率及其对海温变化的响应
|
刘小菊(1997—), 女, 福建省南安市人, 硕士研究生, 主要从事珊瑚礁沉积与环境研究。email: |
Copy editor: 殷波
收稿日期: 2021-11-16
修回日期: 2021-12-28
网络出版日期: 2021-12-29
基金资助
国家科技基础资源调查专项(2018FY100103)
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0206)
中国科学院战略性先导科技专项(A类)(XDA13010103)
The growth rate of coral Porites at the Zhongbei Ansha of the Zhongsha Atoll and its response to the seawater temperature change in the past 165 Years
Copy editor: YIN Bo
Received date: 2021-11-16
Revised date: 2021-12-28
Online published: 2021-12-29
Supported by
National Science & Technology Fundamental Resources Investigation Program of China(2018FY100103)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0206)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA13010103)
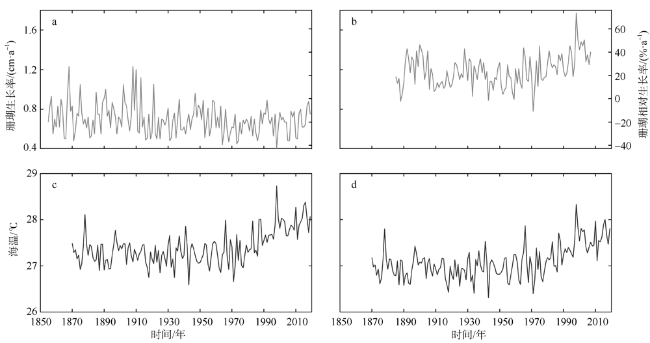
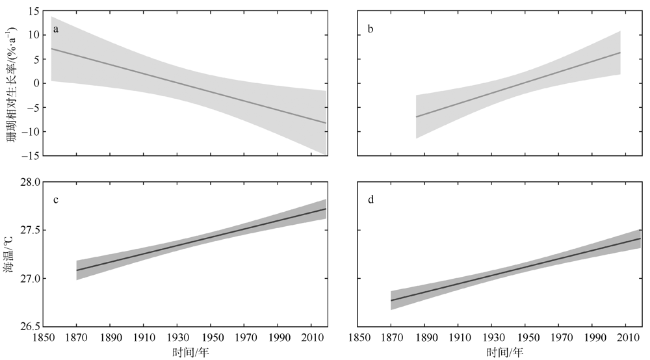
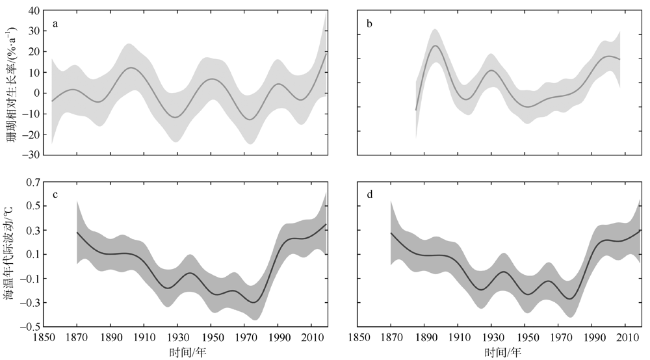
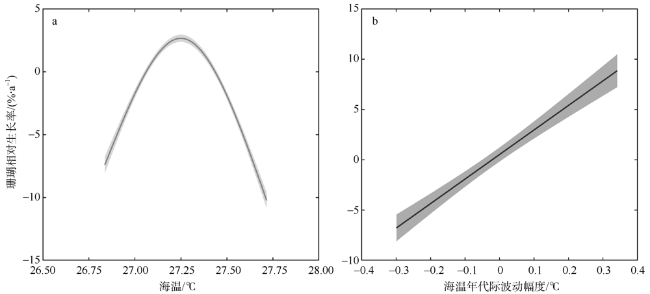
海水温度是控制珊瑚生长的关键环境因素之一, 随着全球气候变暖, 海温持续升高已成为珊瑚生长面临的全球性威胁。文章对采自中沙环礁中北暗沙水深约16m的澄黄滨珊瑚岩心样品开展了生长率分析, 揭示出中沙环礁滨珊瑚近165年来的生长历史及变化规律; 并通过与西沙群岛永兴岛滨珊瑚生长率的对比, 探讨了南海中部滨珊瑚生长的区域差异及其对海温升高的响应关系。过去100多年来中北暗沙和永兴岛海区的平均海温分别为(27.4±0.37)℃和(27.09±0.36)℃, 两个海区的海温均呈线性升高趋势, 升温速率一致, 约为0.43℃·ha-1。过去100多年间中北暗沙和永兴岛滨珊瑚的平均生长率分别为(0.70±0.16)cm·a-1和(1.19±0.16)cm·a-1, 但中北暗沙滨珊瑚生长率呈线性下降趋势, 下降速率约为9.4%·ha-1; 而永兴岛滨珊瑚生长率呈线性上升趋势, 增长速率约为10.9%·ha-1。过去100多年间两个礁区的滨珊瑚生长率均存在年代际波动, 大致与海温的年代际波动对应。两个礁区滨珊瑚生长率与海温在趋势上呈现非线性响应关系, 存在滨珊瑚生长的最适宜温度约为27.25℃, 过去100多年来中北暗沙海域海温的增温趋势已经超出了滨珊瑚生长的适宜海温范围, 限制了滨珊瑚的生长趋势, 而永兴岛海域海温仍适宜滨珊瑚的生长。在年代际波动上两个礁区滨珊瑚生长率与海温存在线性正相关关系, 海温的年代际增温有利于滨珊瑚生长。在南海未来持续海水升温的情况下, 中北暗沙珊瑚生长的下降趋势将会进一步加剧, 并将严重威胁中沙珊瑚礁生态系统的维持和发展。
刘小菊 , 施祺 , 陶士臣 , 杨红强 , 张喜洋 , 周胜男 . 近165年来中沙环礁中北暗沙滨珊瑚生长率及其对海温变化的响应[J]. 热带海洋学报, 2022 , 41(5) : 64 -73 . DOI: 10.11978/2021159
Coral growth is essential for maintaining the coral reef ecosystem and coral reef development. Seawater temperature is one of the key environmental factors to control coral growth. With global warming, the persistent rise of seawater temperature has become a global threat to coral growth. Coral reefs are widely distributed in the South China Sea (SCS), and there is a significant regional difference in coral growth in the SCS. The Zhongsha Atoll, located in the central SCS, is the largest atoll in the SCS. The coral community is well developed, but little is known about the history of coral growth in this reef area. In 2020, we drilled a core sample of coral Porites lutea at water depth of ~16 m at the Zhongbei Ansha of the Zhongsha Atoll. The sample growth rate was analyzed which revealed a 165-year growth history of the coral Porites and its changing pattern at the Zhongbei Ansha. Compared with growth rate of the coral Porites at the Yongxing Island, Xisha Islands, the regional difference of coral growth in the middle part of the SCS and their response to the rise of the sea surface temperature (SST) was discussed. The average SST is about (27.4 ± 0.37) ℃ and (27.09 ± 0.36) ℃, respectively, at the Zhongbei Ansha and the Yongxing Island during the past over a hundred years. SST shows a linearly rising trend at both the two reef areas with a similar rate of about 0.43 ℃·ha-1. The average growth rate of coral Porites is about (0.70 ± 0.16) cm·a-1 and (1.19 ± 0.16) cm·a-1 respectively at the Zhongbei Ansha and the Yongxing Island during the past over a hundred years. The growth rate has a linearly declining trend with a rate of about 9.4 %·ha-1 at the Zhongbei Ansha in the same period, while the growth rate has increased linearly by 10.9 %·ha-1 at the Yongxing Island. In addition, there are interdecadal fluctuations in growth rate in both two areas in the past over a hundred years, approximately corresponding to the interdecadal fluctuations in SST. A nonlinear responding relationship is found between the growth rate of coral Porites and SST within the linear trends at the two areas during the past over a hundred years, and there is an optimum temperature of about 27.25 ℃ for the growth of coral Porites. Moreover, a linear positive correlation exists in the interdecadal fluctuations between growth rate and SST. In the past over a hundred years, seawater warming has exceeded the appropriate temperature range for the growth of coral Porites at the Zhongbei Ansha, limiting the growth trend of coral Porites. At the Yongxing Island, however, the seawater temperature is still suitable for the growth of coral Porites. And then the interdecadal rise of the seawater temperature is conducive to the growth of coral Porites at both areas. Under the condition of continuous seawater warming in the SCS in the future, the declining trend in coral growth will be further aggravated at the Zhongbei Ansha, which will seriously threaten the maintenance and development of the coral reef ecosystem at the Zhongsha Atoll.
Key words: Zhongsha Atoll; Zhongbei Ansha; coral Porites; growth rate; seawater warming
图1 中沙环礁中北暗沙采样点位置(a、b)、滨珊瑚(c)和样品(d)a. 中沙环礁、中北暗沙、西沙群岛和永兴岛。黄色方框处为图b区域; b. 中北暗沙。黄色圆点为采样位置; c. 澄黄滨珊瑚; d. 岩心柱样(左)和切片(右)X光影像 Fig. 1 The location of sampling sites (a, b) and coral Porites (c) and sample (d) from Zhongbei Ansha of Zhongsha Atoll. (a) Zhongsha Atoll, Zhongbei Ansha, Xisha Islands, Yongxing Island; (b) Zhongbei Ansha; (c) Coral Porties lutea; (d) Core sample and slice X-ray image, yellow dot represents the sampling position |
图3 中北暗沙(a、c)和永兴岛(b、d)滨珊瑚相对生长率(a、b)和海温线性趋势(c、d)(阴影为95%置信区间)Fig. 3 Linear trends in the growth rate of coral Porites (a, b) and SST (c, d) at the Zhongbei Ansha (a, c) and the Yongxing Island (b, d) (The shadow area represents a 95 % confidence interval) |
图5 中北暗沙和永兴岛滨珊瑚生长率与海温的响应关系a. 线性趋势响应; b. 年代际波动响应。阴影为95%置信区间 Fig. 5 Responding relationships between the growth rate of coral Porites and SST at the Zhongbei Ansha and Yongxing Island. (a) Response relationship in the linear trends; (b) Response relationship in the interdecadal variations. The shadow area represents a 95 % confidence interval |
| [1] |
樊博文, 雷洁霞, 樊彦国, 2018. 基于GIS的南海海温时空过程分析研究[J]. 海洋科学, 42(4): 36-42.
|
| [2] |
黄博津, 余克服, 张会领, 等, 2013. 利用珊瑚生长率重建西沙海域罗马暖期中期海温变化[J]. 热带地理, 33(3): 237-241.
|
| [3] |
黄金森, 1987. 中沙环礁特征[J]. 海洋地质与第四纪地质, 7(2): 21-24.
|
| [4] |
林丽芳, 余克服, 陶士臣, 等, 2018. 南海黄岩岛珊瑚记录的1780—2013年的表层海温年代际变化特征[J]. 海洋学报, 40(9): 31-42.
|
| [5] |
聂宝符, 陈特固, 粱美桃, 等, 1997. 南沙群岛及其邻近礁区造礁珊瑚与环境变化的关系[M]. 北京: 科学出版社:1-101.
|
| [6] |
聂宝符, 陈特固, 彭子成, 1999. 由造礁珊瑚重建南海西沙海区近220a海面温度序列[J]. 科学通报, 44(17): 1885-1889.
|
| [7] |
牛明星, 2018. 1871—2010年南海及毗邻的西北太平洋海域盐度场的时空变化特征[D]. 上海: 上海师范大学:12-32.
|
| [8] |
施祺, 余克服, 陈天然, 等, 2012. 南海南部美济礁200余年滨珊瑚骨骼钙化率变化及其与大气CO2和海水温度的响应关系[J]. 中国科学 D辑: 地球科学, 42(1): 71-82.
|
| [9] |
谭红建, 蔡榕硕, 颜秀花, 2016. 基于IPCC-CMIP5预估21世纪中国近海海表温度变化[J]. 应用海洋学学报, 35(4): 451-458.
|
| [10] |
陶士臣, 张会领, 余克服, 等, 2021. 近500年西沙群岛海面温度年际变化的珊瑚记录及其环境意义[J]. 第四纪研究, 41(2): 411-423.
|
| [11] |
佟飞, 陈丕茂, 秦传新, 等, 2015. 南海中沙群岛两海域造礁石珊瑚物种多样性与分布特点[J]. 应用海洋学学报, 34(4): 535-541.
|
| [12] |
张会领, 余克服, 施祺, 等, 2014. 珊瑚生长率重建西沙海域中晚全新世海温变化[J]. 第四纪研究, 34(6): 1296-1305.
|
| [13] |
邹仁林, 2001. 中国动物志腔肠动物门珊瑚虫纲石珊瑚目造礁石珊瑚[M]. 北京: 科学出版社: 1-248.
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}