
海南澳洲管体星虫卵细胞发育及生殖周期研究
|
乔立君(1998—), 男, 山东省济宁市人, 在读硕士研究生, 从事海洋生物学方面研究。email: |
Copy editor: 殷波
收稿日期: 2021-10-25
修回日期: 2021-12-28
网络出版日期: 2021-12-31
基金资助
海南省自然科学基金(319MS013)
Development of oocytes and reproductive cycle of Siphonosoma australe in Hainan
Copy editor: YIN Bo
Received date: 2021-10-25
Revised date: 2021-12-28
Online published: 2021-12-31
Supported by
Hainan Provincial Natural Science Foundation of China(319MS013)
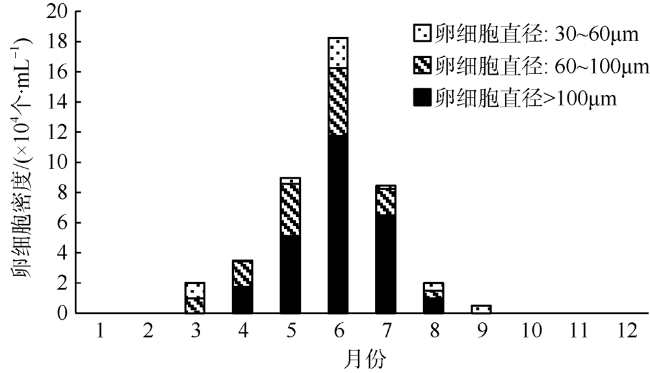
澳洲管体星虫(Siphonosoma australe)是海南当地极具特色的星虫资源, 经济价值高。近年来该资源急剧衰竭, 开展星虫资源恢复与保护十分重要。文章对海南澳洲管体星虫卵细胞的显微与超微结构进行研究。结果显示: 1) 卵细胞发育经历4个阶段: 生长前期, 体腔液中大量光滑的凹饼状红细胞出现表面凹凸不平的褶皱(直径小于30μm), 暗示这些红细胞是卵原细胞; 生长后期, 30~40μm的卵细胞表面褶皱消失, 变成光滑的圆厚饼状, 并出现厚度1μm的卵黄膜, 卵黄颗粒开始积累; 成熟前期, 卵细胞球状, 直径60~120μm, 卵黄膜增厚至5~11μm, 卵黄颗粒增多, 细胞核增大, 部分染色质形成高电子密度的团块散布于核内; 成熟后期, 直径120μm, 卵黄膜厚11~12μm, 卵黄颗粒充满整个卵细胞, 膜孔外露于膜表层。2) 成熟期澳洲管体星虫卵黄膜仅分为两层: 有一定厚度的均质内层和多层重叠膜结构组成的外层, 同时外层上覆盖有粒状突。膜孔结构简单, 主要是膜凹陷折叠形成。3) 卵黄颗粒分为Ⅰ型和Ⅱ型, 其发生途径主要由线粒体、内质网、高尔基体和溶酶体等细胞器演变而成, 亦可由卵母细胞吞饮而成。最为常见的是高尔基体包裹形成的Ⅱ型卵黄颗粒。4) 海南文昌海域澳洲管体星虫的繁殖季节为4—8月, 其中5—7月是繁殖高峰期。3月初卵母细胞开始出现, 4—8月成熟卵细胞在体腔中保持一定密度, 9月份卵细胞密度降至极低, 10月份后消失。澳洲管体星虫作为一种新的具有热带特色的水产种质资源, 其卵细胞发育及生殖周期研究将极大推动其繁育及保护的相关技术探索。
乔立君 , 姚雪梅 , 余巧驰 . 海南澳洲管体星虫卵细胞发育及生殖周期研究[J]. 热带海洋学报, 2022 , 41(5) : 161 -169 . DOI: 10.11978/2021143
Siphonosoma australe is an important sipuculan species and a typical local resource in Hainan with medicinal and edible value, it is distributed in tropical and subtropical coastal waters, mainly in surrounding waters of Hainan island. Due to overfishing and habitat destruction, the population size of S. australe rapidly declined in recent years. So the recovery and protection of the sipuculan resource is very important. The structures of oocytes of S. australe were observed by optical microscope and electron microscope. The results are shown as follows: 1) The oogenesis of S. australe could be divided into four stages: In the early growth stage, concave disk-shaped coelomocytes (less than 30 μm in diameter) changed from the smooth cells to lumpy and corrugated cells, which suggested that concave coelomocytes were the oogonia; in the late growth stage, the surface of 30 ~ 40 μm oocytes became smooth and unfolded, and the vitelline membrane of 1 μm in thickness appeared with the accumulation of yolk granules; in the early mature stage, oocytes were 60 ~ 120 μm in diameter and transformed into spherical shape. The yolk granules increased. The nucleus was enlarged. Some of the chromatin formed clumps with high electron density and dispersed in the nucleus; in the late mature stage, oocytes were 120 μm in diameter with the yolk granules filling the whole cells, and the membrane pores were exposed on the membrane surface. 2) The vitelline membrane of mature oocyte can be divided into two layers: the homogeneous inner layer with a thin thickness and the multi-folded outer layer with granular protuberances. The structure of membrane micropore was simple and formed by the invagination of the folded membranes. 3) During oogenesis, a variety of organelles increased and the yolk granules (type Ⅰ and Ⅱ) were combined with organelles, e.g. mitochondria, endoplasmic reticulum, Golgi apparatus, lysosome, evenly transformed by phagocytic vesicle. The type Ⅱ yolk-granules enveloped by Golgiosomes were common. 4) The reproductive season of S. australe in Wenchang, Hainan is from April to August and the reproductive peak is in May and July. Oocytes begin to appear in early March, quantitatively drop to a very low level in September and disappear in October. The mature oocytes keep a certain density in the coelom from April to August. It is speculated that the reproduction period is related to the monthly water-temperature and the season of highest water-temperature is the period of reproductive peak. As a new Hainan fishery resource with tropical characteristics, this research on oocyte development and reproductive cycle of S. australe is expected to greatly promote the development on related technologies for reproduction and protection.
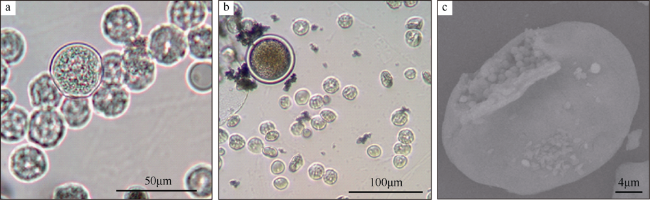
图1 澳洲管体星虫卵细胞生长前期a. 表面光滑的红细胞(smooth erythrocyte, SE)和表面凹凸不平的饼状卵母细胞(uneven surface erythrocyte, USE); b和c. 表面光滑的红细胞(SEM/TEM); d. 表面凹凸不平的饼状细胞(SEM); e. 双面凹陷的表面凹凸不平的卵母细胞(TEM); f. 表面凹凸不平的卵母细胞(TEM), V表示囊泡 Fig. 1 The oocytes in the early growth stage of Siphonosoma australe. (a) smooth erythrocyte (SE) and uneven surface erythrocyte (USE); (b) and (c) red blood cell with a smooth surface (SEM/TEM); (d) disk-like cells with the uneven surfaces (SEM); (e) double-side concave oocyte with vesiculose, lumpy and corrugated surface (TEM); (f) oocyte with an uneven surface (TEM). V: vesicle |
图3 澳洲管体星虫卵细胞成熟前期a. 卵细胞球形, 膜孔被表面膜覆盖(SEM); b. 凹陷的膜孔被表面膜覆盖和卵黄膜的内外两层(TEM); c~e. 各种卵黄颗粒形态和大量细胞器(TEM); f. 核膜周围大量出现的内质网(TEM); g. 被高尔基体包围的Ⅱ型卵黄颗粒(TEM); h. 染色质形成高电子密度团块(TEM)。图中V1表示卵黄膜外层, V2表示卵黄膜内层, P表示粒状突, MPV表示胞饮泡, L表示溶酶体, G表示高尔基体, M表示线粒体, ER表示内质网, Y1表示Ⅰ型卵黄颗粒, Y2表示Ⅱ型卵黄颗粒, MB表示髓样小体, N表示细胞核, Ch表示染色质 Fig. 3 The oocytes in the early mature stage of Siphonosoma australe. (a) spherical oocyte, membrane pores masked by a superficial vitelline membrane (SEM); (b) sunken membrane-pores and two layers (V1 and V2) forming the vitelline membrane; (c) ~ (e) morphology of a variety of yolk particles and numerous organelles (TEM); (f) numerous endoplasmic reticulum appearing around the nuclear membrane (TEM); (g) yolk particles surrounded by the Golgi (TEM); (h) chromatin forming clumps with high electron density (TEM). V1: outer layer of vitelline membrane; V2: inner layer of vitelline membrane; P: granular protuberance; MPV: micropinocytotic vesicle; L: lysosome; G: Golgi body; M: mitochondria; ER: endoplasmic reticulum; Y1: type I yolk granule; Y2: type II yolk granule; MB: myeloid body; Ch: chromatin |
图4 澳洲管体星虫卵细胞成熟后期a. 成熟卵细胞(直径120μm); b. 外膜部分剥落(SEM); c. 破膜后可见膜孔(SEM); d. 卵黄膜上的胞饮泡(TEM); e. 大量的Ⅱ型卵黄颗粒, 主要由大量的高尔基体包裹的(TEM); f. 卵黄颗粒填满整个卵细胞(TEM)。图中PM表示放大的膜孔结构, Y2表示高尔基体包裹形成的卵黄颗粒, N表示细胞核 Fig. 4 The oocytes in the late mature stage of Siphonosoma australe. (a) mature oocytes (120 μm in diameter); (b) exfoliation of the outer-layer membrane (SEM); (c) clear visible micropores after peeling of outermost membrane; (d) micropinocytotic vesicles in the vitelline membrane; (e) large number of yolk particles, mainly contained by the Golgi bodies; (f) yolk granules filling to the cytoplasm of a mature oocyte. PM: magnified membrane-pore structure, Y2: type II yolk particles formed by Golgi wrapping, N: nucleus |
| [1] |
陈慧, 林国文, 陈武, 等, 2009. 可口革囊星虫生殖周期的观察[J]. 海洋渔业, 31(2): 139-145.
|
| [2] |
陈子安, 杜晓东, 王庆恒, 等, 2007. 3种星虫线粒体16S rRNA、CO I和Cyt b基因片段的序列比较[J]. 广东海洋大学学报, 27(4): 3-10.
|
| [3] |
顾晓英, 竺俊全, 许式见, 等, 2009. 可口革囊星虫(Phascolosoma esculenta)卵子发生的组织学研究[J]. 海洋与湖沼, 40(3): 283-288.
|
| [4] |
郭学武, 李复雪, 1993. 光裸星虫生殖周期的研究[J]. 热带海洋学报, 12(2): 69-76.
|
| [5] |
兰国宝, 阎冰, 2002. 方格星虫繁殖生物学研究[J]. 水产学报, 26(6): 503-509.
|
| [6] |
李凤鲁, 周红, 王玮, 1992. 中国沿海管体星虫属(星虫动物门)的研究[J]. 青岛海洋大学学报, 22(1): 97-102.
|
| [7] |
刘懂, 龙玲利, 曾海祥, 等, 2014. 可口革囊星虫体腔中卵细胞大小组成的周年变化[J]. 生物学杂志, 31(4): 29-32.
|
| [8] |
王庆恒, 杜晓东, 黄洪艳, 等, 2005. 湛江地区光裸星虫的生殖细胞发育和生殖周期[J]. 湛江海洋大学学报, 25(1): 5-9.
|
| [9] |
王庆恒, 张家炜, 郝瑞娟, 等, 2017. 光裸星虫体腔液中卵子发生的超微结构[J]. 海洋与湖沼, 48(1): 57-66.
|
| [10] |
吴斌, 1999. 光裸方格星虫(Sipunculus nudus L.)生殖细胞及胚胎发育[J]. 广西科学, 6(3): 222-226.
|
| [11] |
徐海娇, 2012. AFM研究高尔基体和内质网的结构[D]. 长春: 东北师范大学.
|
| [12] |
周红, 李凤鲁, 王玮, 2007. 中国动物志无脊椎动物第四十六卷星虫动物门螠虫动物门[M]. 北京: 科学出版社.
|
| [13] |
竺俊全, 王伟, 丁理法, 2012. 可口革囊星虫(Phascolosoma esculenta)卵黄合成期卵母细胞发育及卵黄发生与卵膜形成的超微结构[J]. 海洋与湖沼, 43(4): 870-876.
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
/
| 〈 |
|
〉 |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}