
马氏珠母贝异速生长个体对免疫刺激的差异响应
|
许瀚之(1995—), 男, 山东省高密市人, 硕士研究生, 从事马氏珠母贝分子遗传育种研究。email: |
Copy editor: 殷波
收稿日期: 2021-07-27
修回日期: 2021-12-31
网络出版日期: 2022-01-12
基金资助
国家贝类产业技术体系项目(CARS-49)
国家自然科学基金青年科学基金(42006106)
广东省自然科学基金(2020A1515011434)
广东省科技计划(2020B1212060058)
Differential responses of allometric individuals to immune stimuli in Pinctada fucata martensii
Copy editor: YIN Bo
Received date: 2021-07-27
Revised date: 2021-12-31
Online published: 2022-01-12
Supported by
National Shellfish Industry Technology System Project(CARS-49)
National Natural Science Foundation of China(42006106)
Natural Science Foundation of Guangdong Province(2020A1515011434)
Science and Technology Program of Guangdong Province(2020B1212060058)
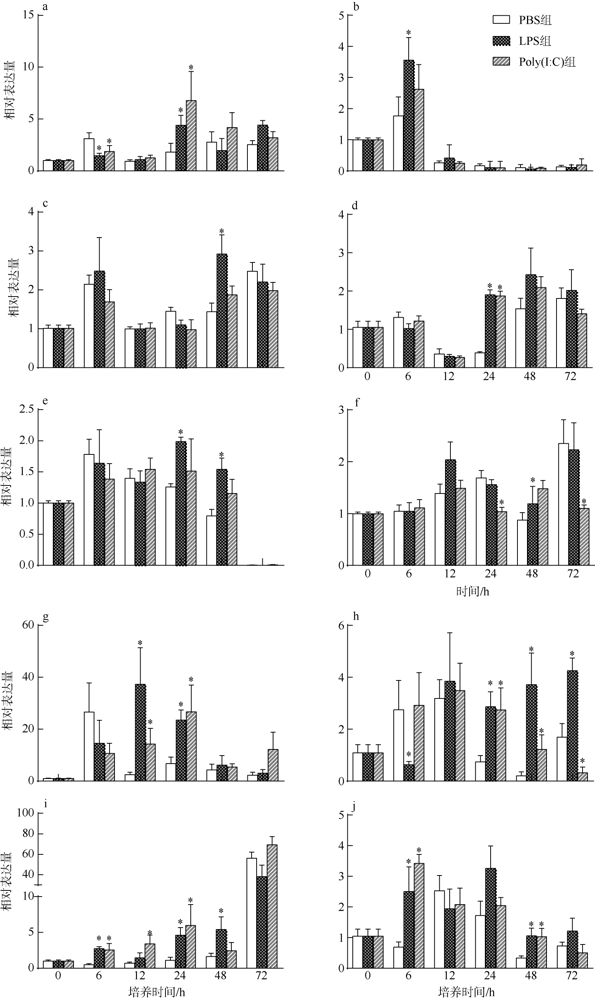
文章对马氏珠母贝的异速生长个体的免疫性能进行了研究, 比较了极大和极小个体在免疫刺激后免疫相关基因(CuZn-SOD1、CuZn-SDO2、LAMP1、JRL和Rab7)的表达量以及3种免疫酶(SOD、CAT、AKP)活性的变化情况, 分析异速生长个体对免疫刺激的差异响应。结果显示, 所检测的与免疫相关的基因和免疫酶对免疫刺激均有应答。极小个体的LAMP1、CuZn-SOD1和CuZn-SOD2基因的相对表达量以及CAT和AKP活性的变化幅度均高于极大个体, 而JRL和Rab7基因的相对表达量以及SOD活性的变化幅度低于极大个体, 其中极小个体JRL基因的基础表达量远高于极大个体, 可能导致其变化幅度较小。研究结果表明, 极小个体对外界胁迫的反应更加强烈, 具有较强的免疫响应能力。
许瀚之 , 张华 , 熊盼盼 , 何毛贤 . 马氏珠母贝异速生长个体对免疫刺激的差异响应[J]. 热带海洋学报, 2022 , 41(5) : 180 -188 . DOI: 10.11978/2021095
In this study, we investigated the immune performance of allometric individuals of the pearl oyster, Pinctada fucata martensii, compared the expression of immune-related genes (CuZn-SOD1, CuZn-SDO2, LAMP1, JRL, and Rab7) and changes in the activities of three immune enzymes (SOD, AKP and CAT) in large and small individuals after immune stimulation, and analyzed the differential response of allometric individuals to immune stimulation. The results showed that the immune-related genes and immune enzymes all responded to the immune stimuli. The relative expression levels of LAMP1, CuZn-SOD1 and CuZn-SOD2 genes and the changes of CAT and AKP activities were higher in small individuals than those in large individuals, while the relative expression levels of JRL and Rab7 genes and the changes of SOD activity were lower than those in large individuals. The basal expression of the JRL gene was much higher in the small individuals than that in the large ones, which may result in a smaller variation. These results indicated that the small individuals were more responsive to external stresses and had stronger immune response ability.
表1 实时荧光定量引物Tab. 1 Real-time quantitative primers |
| 引物名称 | 引物序列 |
|---|---|
| CuZn-SOD1-F | ACAAATAATTCGGCATCCTA |
| CuZn-SOD1-R | TAGAGCCTGGTTTGGTGTAT |
| CuZn-SOD2-F | ATAGGTTCAAACGGAGGAGG |
| CuZn-SOD2-R | TGGTCGCAAATGGTTGTATT |
| JRL-F | ATAGGTTCAAACGGAGGAGG |
| JRL-R | TGGTCGCAAATGGTTGTATT |
| LAMP-1F | TGCCTGTCACAATAATAACC |
| LAMP1-R | AGACTCAAAGTAAGACCACCT |
| Rab7-F | GAGTTTAGGCGTGGCTTTCT |
| Rab7-R | ATCCCTTGGACTTGCTTGTAT |
| LAMP2-F | ACGAAAAAAAGGTTTGAGAGACG |
| LAMP2-R | AGATACTTGATGGCCTGTTG |
| 18S-F | CGTTTCAACAAGACGCCAGTAG |
| 18S-R | ACGAAAAAAAGGTTTGAGAGACG |
| [1] |
陈丹群, 2009. 合浦珠母贝家系选育及免疫酶和消化酶活性研究[D]. 武汉: 华中农业大学.
|
| [2] |
刘婷婷, 齐洪庆, 李月, 等, 2019. 仿刺参过氧化氢酶的克隆、表达及纯化[J]. 大连工业大学学报, 38(6): 398-402.
|
| [3] |
牟海津, 江晓路, 刘树青, 等, 1999. 免疫多糖对栉孔扇贝酸性磷酸酶、碱性磷酸酶和超氧化物歧化酶活性的影响[J]. 青岛海洋大学学报, 29(3): 463-468.
|
| [4] |
孙宗红, 刘志刚, 白成, 等, 2015. 温度和盐度对4种壳色马氏珠母贝谷胱甘肽过氧化物酶基因表达的影响[J]. 广东海洋大学学报, 35(4): 24-31.
|
| [5] |
王哲, 王姿曼, 郝瑞娟, 等, 2019. 马氏珠母贝金黄壳色选育群体植核后差异表达免疫基因筛选[J]. 广东海洋大学学报, 39(6): 9-16.
|
| [6] |
魏海军, 邓正华, 陈明强, 等, 2020. 不同壳色合浦珠母贝生产性能评估及其雌雄群体间的差异[J]. 南方农业学报, 51(2): 412-420.
|
| [7] |
张志东, 张雨, 陈爱华, 等, 2019. 文蛤过氧化氢酶的生物信息学分析[J]. 生物信息学, 17(4): 249-255.
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}