
桐花树红树林潮滩近底层悬沙浓度垂向剖面变化特征分析*
|
黄祖明(1997—), 硕士研究生, 主要从事河口海岸动力地貌研究。email: |
Copy editor: 姚衍桃
收稿日期: 2021-12-01
修回日期: 2022-01-26
网络出版日期: 2022-02-06
基金资助
国家自然科学基金重点项目(41930537)
广西重点研发计划(桂科)(AB21076016)
Analysis of the variations of suspended sediment concentration in vertical profile near the bed of Aegiceras corniculatum tidal flat
Copy editor: YAO Yantao
Received date: 2021-12-01
Revised date: 2022-01-26
Online published: 2022-02-06
Supported by
National Natural Science Foundation of China(41930537)
Guangxi Key Research and Development Program(AB21076016)
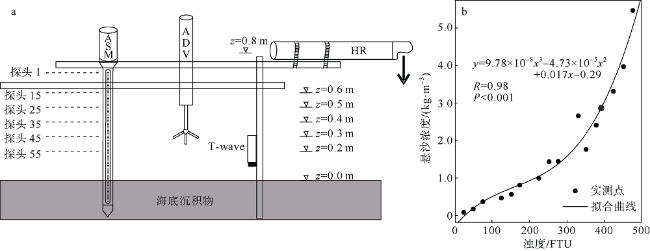
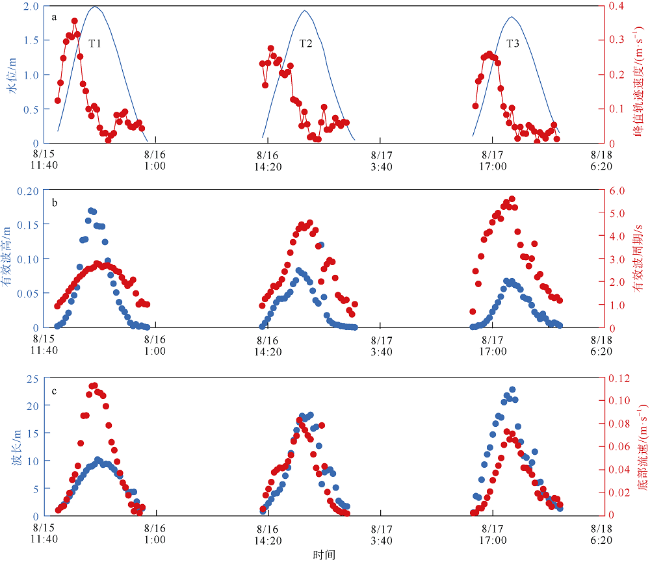
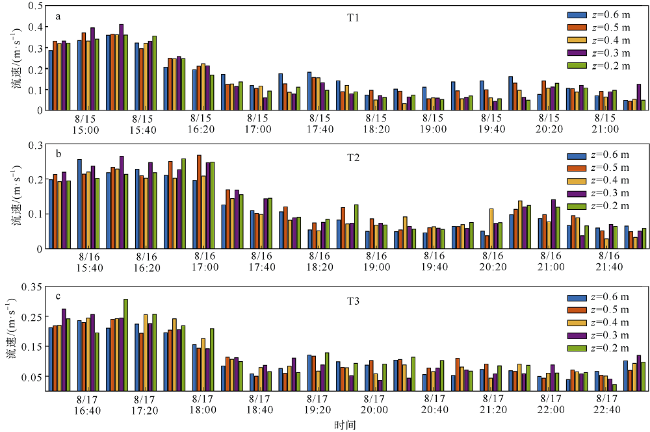
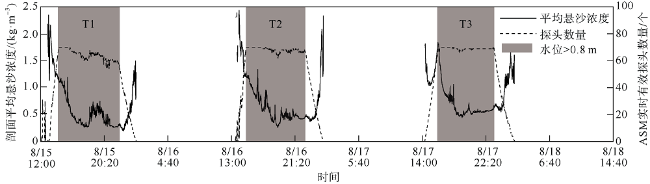
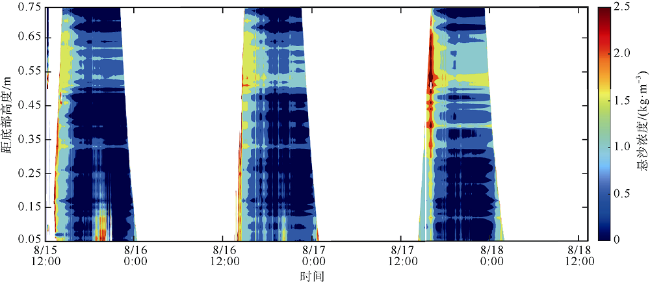
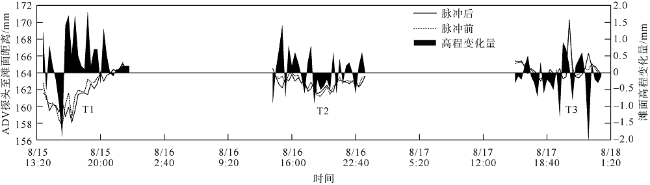
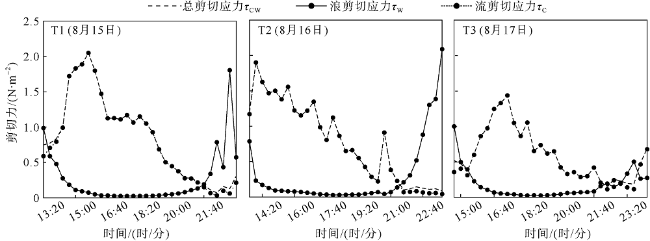
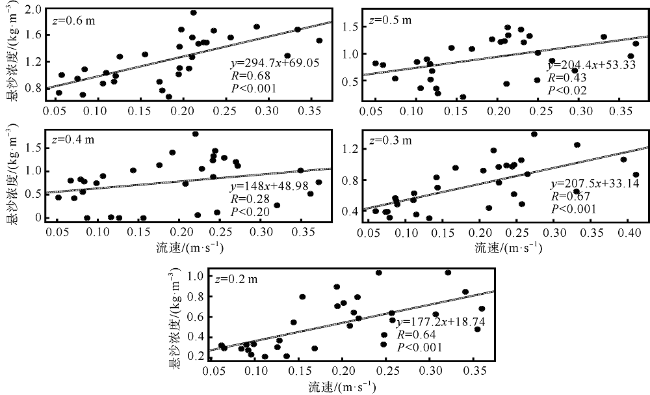
观测红树林潮滩在波浪和潮流作用下的近底层垂向剖面悬沙浓度变化过程, 对理解海岸带植被的消能促淤机制和滨海湿地生态修复工程有着重要作用。本文以北部湾七星岛岛尾桐花树红树林潮滩为例, 基于剖面流速仪HR、声学多普勒单点流速仪ADV、浪潮仪T-wave及剖面浊度仪ASM, 获取了研究区域2019年夏季大潮连续3天的水文数据, 同时结合桐花树典型植株实测参数, 分析了潮周期内红树林潮滩近底层垂向剖面悬沙响应波浪、潮流作用及桐花树空间结构的运动过程。结果表明: 1) 桐花树潮滩近底层悬沙浓度和悬沙通量具有涨潮明显大于落潮的潮汐不对称现象, 剖面垂向高悬沙浓度区域在涨潮初期—涨急由距底部0.1~0.37m转变为距底部0.5~0.67m, 落急—落潮末期则由上部转变为下部; 2) 潮周期内悬沙起动和再悬浮阶段发生在以波浪作用主导的涨潮初期和落潮末期, 平流和沉降发生在以潮流作用为主的涨急至落急整个阶段; 3) 涨潮阶段桐花树冠层的茂密枝叶通过减缓流速拦截多于冠层上部40%以上的悬沙, 落潮水体则挟沙自陆向海经过桐花树群落, 使得悬沙浓度下降超过71%。该不对称涨、落潮动力沉积机制有利于悬沙向岸输运, 促进潮滩扩张过程。
黄祖明 , 周晓妍 , 戴志军 , 车志伟 . 桐花树红树林潮滩近底层悬沙浓度垂向剖面变化特征分析*[J]. 热带海洋学报, 2022 , 41(4) : 38 -50 . DOI: 10.11978/2021167
Suspended sediment concentration(SSC)observation in vertical profile near the bed of mangrove tidal flat under the waves and tidal currents, is important to understand the mechanisms of energy dissipation and deposition promotion by vegetations, and the ecological restoration in coastal zone. In this study, we measured the hydrological data using HR, ADV, Twave and ASM and the typical plants’ parameters of Aegiceras corniculatum for 3 consecutive days of flood tides in the summer of 2019. Then the process of the near-bed vertical SSC of mangrove tidal flat was analyzed in response to waves, tidal currents and spatial structure of Aegiceras corniculatum. The results showed that: (1) the variations of suspended sediment flux and SSC near bed in Aegiceras corniculatum tidal flat had the characteristic of tidal asymmetry, the SSC of flood tide was significantly greater than that of ebb tide. The vertical high SSC area of the profile changed from 0.1~0.37 m to 0.5-0.67 m from the bottom in the early flood tide to flood maximum, and from the upper to the bottom in ebb maximum to end. (2) In the intra-tide, the suspension and resuspension stages occurred at the early flood and latest ebb that were dominated by waves. The advection and deposition stages occurred at the whole period from flood maximum to ebb maximum dominated by tidal currents. (3) Dense branches and leaves in the canopy of Aegiceras corniculatum intercepted more than 40% of the suspended sediment in the upper canopy during flood tide. The SSC decreased by more than 71% when the flow during ebb tide. This dynamical deposition mechanism of asymmetrical flood and ebb tide was conducive to sediment transportation landward which then led to flat expansion.
表1 近底层剖面各高度在涨、落潮期间的悬沙通量Tab. 1 Suspended sediment flux per unit area at each height in profile near bed |
| 潮周期 | 悬沙通量/(kg·m-3) | |||||
|---|---|---|---|---|---|---|
| z=0.6m | z=0.5m | z=0.4m | z=0.3m | z=0.2m | ||
| T1 | 涨潮 | 3.013 | 1.737 | 0.839 | 1.721 | 1.154 |
| 落潮 | -0.893 | -0.130 | -0.002 | -0.219 | -0.303 | |
| T2 | 涨潮 | 2.382 | 2.126 | 1.686 | 1.672 | 1.124 |
| 落潮 | -0.676 | -0.528 | -0.374 | -0.278 | -0.176 | |
| T3 | 涨潮 | 2.321 | 2.016 | 2.231 | 1.669 | 1.049 |
| 落潮 | -0.898 | -0.807 | -0.714 | -0.383 | -0.219 | |
表2 七星岛岛尾研究区域典型植株参数平均值Tab. 2 Average values of typical Aegiceras corniculatum parameters in study area |
| 植株参数 | 典型中株 | 典型小株 |
|---|---|---|
| 数目/棵 | 26 | 8 |
| 高度/cm | 66 | 17 |
| 冠层长度/cm | 85 | 15 |
| 冠层宽度/cm | 45 | 11 |
| Dz=0.1/cm | 1.6 | 0.5 |
| Dz=0.3/cm | 0.8 | |
| Dz=0.5/cm | 0.4 | |
| 树枝总数目/条 | 72 | 24 |
| 树叶总数目/片 | 210 | 75 |
| [1] |
戴志军, 周晓妍, 王杰, 等, 2021. 红树林潮滩沉积动力研究进展与展望[J]. 热带海洋学报, 40(3): 69-75.
|
| [2] |
蒋磊明, 陈波, 邱绍芳, 2008. 廉州湾三角洲泥沙运移与海洋动力条件的关系[J]. 广西科学院学报, 24(1): 25-28.
|
| [3] |
江锐捷, 程鹏, 高建华, 等, 2020. 红树林对潮流底边界层动力过程的影响[J]. 海洋地质前沿, 36(4): 37-44.
|
| [4] |
李春干, 2003. 广西红树林资源的分布特点和林分结构特征[J]. 南京林业大学学报(自然科学版), 27(5): 15-19.
|
| [5] |
李华, 杨世伦, 2007. 潮间带盐沼植物对海岸沉积动力过程影响的研究进展[J]. 地球科学进展, 22(6): 583-591.
|
| [6] |
李华, 杨世伦, 2010. 潮间带盐沼植物黏附悬浮颗粒物的差异性研究[J]. 海洋学报, 32(1): 114-119.
|
| [7] |
林镇坤, 王爱军, 叶翔, 2019. 南流江河口水下三角洲表层沉积物端元分析及其沉积动力环境意义[J]. 沉积学报, 37(1): 124-134.
|
| [8] |
莫永杰, 1988. 南流江河口动力过程与地貌发育[J]. 海洋通报, 7(3): 41-46. (in Chinese)
|
| [9] |
王日明, 戴志军, 黄鹄, 等, 2021. 南流江河口桐花树生物动力地貌过程研究[J]. 海洋学报, 43(9): 102-114.
|
| [10] |
汪亚平, 高抒, 贾建军, 2006. 浪流联合作用下潮滩沉积动力过程的高分辨率数据采集与分析[J]. 科学通报, 51(3): 339-348.
|
| [11] |
杨世伦, 陈吉余, 1994. 试论植物在潮滩发育演变中的作用[J]. 海洋与湖沼, 25(6): 631-635.
|
| [12] |
张乔民, 温孝胜, 宋朝景, 等, 1996. 红树林潮滩沉积速率测量与研究[J]. 热带海洋, 15(4): 57-62.
|
| [13] |
赵焕庭, 张乔民, 宋朝景, 等, 1999. 华南海岸和南海诸岛地貌与环境[M]. 北京: 科学出版社. (in Chinese)
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
/
| 〈 |
|
〉 |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}