
三亚湾珊瑚来源虫黄藻不同株系微环境中微生物群落结构的差异比较分析
|
黄思军(1982—), 男, 福建省光泽县人, 项目研究员, 从事海洋微生物生态学研究。email: |
Copy editor: 林强
收稿日期: 2022-01-23
修回日期: 2022-03-29
网络出版日期: 2022-04-12
基金资助
国家自然科学基金项目(41576126)
国家自然科学基金项目(42176116)
广东省自然科学基金(2017A030306020)
中国科学院南海海洋研究所“南海新星”项目(NHXX2019ST0101)
中国科学院青年创新促进会会员(2018377)
Phycosphere microbial communities of zooxanthellae cultures isolated from corals in Sanya Bay, South China Sea
Copy editor: LIN Qiang
Received date: 2022-01-23
Revised date: 2022-03-29
Online published: 2022-04-12
Supported by
National Natural Science Foundation of China(41576126)
National Natural Science Foundation of China(42176116)
Natural Science Foundation of Guangdong Province(2017A030306020)
Rising Star Foundation of the South China Sea Institute of Oceanology(NHXX2019ST0101)
Youth Innovation Promotion Association CAS(2018377)
虫黄藻、细菌和造礁石珊瑚有着密切的共生关系, 但虫黄藻藻际细菌群落尚未得到广泛研究。本研究对5个属的6株虫黄藻进行了离体培养, 其中2株为悬浮型虫黄藻(E型), 4株为贴壁型虫黄藻(A—D型)。通过采集藻株培养物3种粒径的样本开展细菌群落分析, 分别为0.2~3μm(自由生活)、>3μm(附着于藻体或颗粒物)与Settling(沉底贴壁藻体上)。结果发现, 2种生活方式的藻株藻际细菌群落具有显著差别, 贴壁型藻株细菌群落的物种丰富度显著高于悬浮型藻株。发现7个属的细菌广泛存在于所有的藻株中, 它们代表了A—E型虫黄藻藻际细菌的核心类群。对3种粒径样本的核心细菌群落比较发现, 自由生和颗粒附着生的核心细菌群落十分相似, 但均与沉底贴壁样本差异明显。
黄思军 , 邱晨 , 龙超 , 龙丽娟 . 三亚湾珊瑚来源虫黄藻不同株系微环境中微生物群落结构的差异比较分析[J]. 热带海洋学报, 2022 , 41(6) : 90 -104 . DOI: 10.11978/2022013
Zooxanthellae, bacteria, and reef—building corals have a close symbiotic relationship, but the phycosphere bacterial community of zooxanthellae is poorly studied. In this study, six strains of zooxanthellae from five genera were cultured in vitro, among which two were free-living zooxanthellae strains (type E) and four were anchorage-dependent living zooxanthellae strains (type A—D). Bacterial community analysis was performed by collecting three different particle size fractions (0.2-3μm, >3μm, and settling algae cells). Results showed that the phycosphere bacterial communities between the two lifestyle algal strains were significantly different. The bacterial species richness in anchorage-dependent living strains was significantly higher than that in free-living strains. Seven bacterial genera were observed to be present in all the algal strains, representing the core bacterial communities of zooxanthellae. Comparing the core bacterial communities in three particle size samples, we found that the core bacterial communities in >3μm samples and in 0.2-3μm samples are highly similar, but both were significantly different from the “Settling” samples.
表1 虫黄藻藻株来源及分类信息Tab. 1 Source and taxonomic information of zooxanthellae strains |
| 藻株 | 宿主拉丁文名(中文名) | ITS2基因型 | 藻株分类(属级) |
|---|---|---|---|
| SYSC-2-1 | Acropora tenuis (柔枝鹿角珊瑚) | C1 | Cladocopium (C型) |
| SYSC-2-8 | Acropora tenuis (柔枝鹿角珊瑚) | E101 | Effrenium (E型) |
| SYSC-14-11 | Galaxea fascicularis (丛生盔形珊瑚) | E101 | Effrenium (E型) |
| SYSC-17-3 | Galaxea fascicularis (丛生盔形珊瑚) | A6 | Symbiodinium (A型) |
| SYSC-24-3 | Platygyra verwyi (小叶扁脑珊瑚) | D1 | Durusdinium (D型) |
| SYSC-28-9 | Pavona decussate (十字牡丹珊瑚) | B1 | Breviolum (B型) |
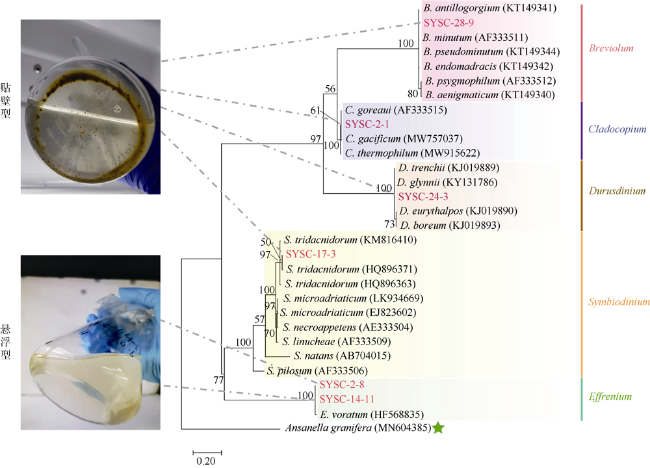
图1 基于ITS2序列构建的系统发育树(ML法)本实验的6株虫黄藻用红色字体标注。Ansanella granifera为外类群。支上的数字为1000次自展支持率(%)。6株虫黄藻被分为贴壁型(SYSC-2-1, SYSC-17-3, SYSC-24-3, SYSC-28-9)和悬浮型(SYSC-2-8和SYSC-14-11) Fig. 1 Maximum-likelihood phylogenetic tree based on ITS2 sequences. The six strains of zooxanthellae in this experiment are marked in red. Ansanella granifera is used as the outgroup. The bootstrap value (%) with 1000 replicates is shown above the branch. The six zooxanthellae strains are classified into anchorage-dependent living lifestyle (SYSC-2-1, SYSC-17-3, SYSC-24-3, SYSC-28-9) and free-living lifestyle (SYSC-2-8 and SYSC-14-11), as indicated in the pictures |
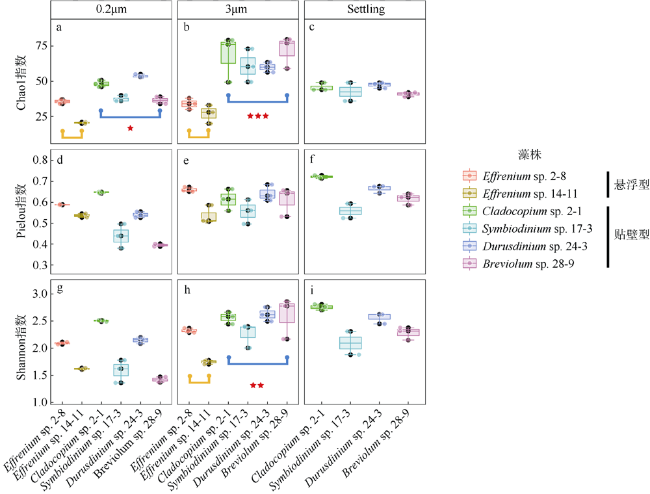
图2 6株虫黄藻藻际细菌群落的Alpha多样性(Chao1、Pielou、Shannon指数)包括所有虫黄藻藻株的3μm和0.2μm样本, 及4株贴壁型藻的Settling样本。显著性标记***表示P<0.001; **表示P<0.01; *表示P<0.05 Fig. 2 The α diversity indices (Chao1, Shannon, Pielou) of phycosphere bacterial communities in six zooxanthellae strains, including 3 μm and 0.2 μm samples of all zooxanthellae strains, as well as “Settling” samples of four anchorage-dependent living algae. Significance code: ***, P < 0.001; **, P < 0.01; *, P < 0.05 |
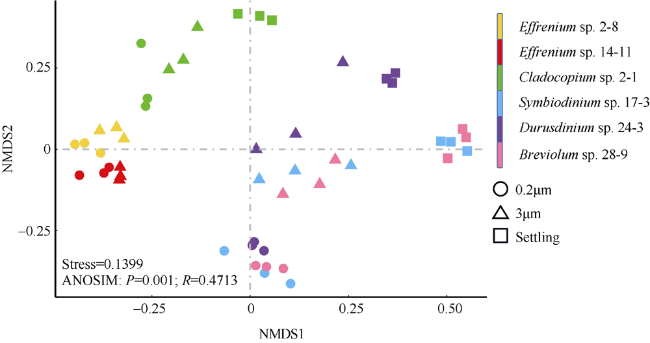
图3 基于Bray-Curtis相异指数的虫黄藻藻际细菌群落的非度量多维标度(NMDS)分析ANOSIM分析指示6株虫黄藻藻际细菌群落的整体差异 Fig. 3 Non-metric multidimensional scaling (NMDS) analysis based on the Bray-Curtis dissimilarity among bacterial community of zooxanthellae. Three types of samples are represented by different shapes (circles for 0.2μm samples, triangles for 3μm samples, and squares for Settling samples). The ANOSIM analysis indicates the overall similarity among the six algal strains |
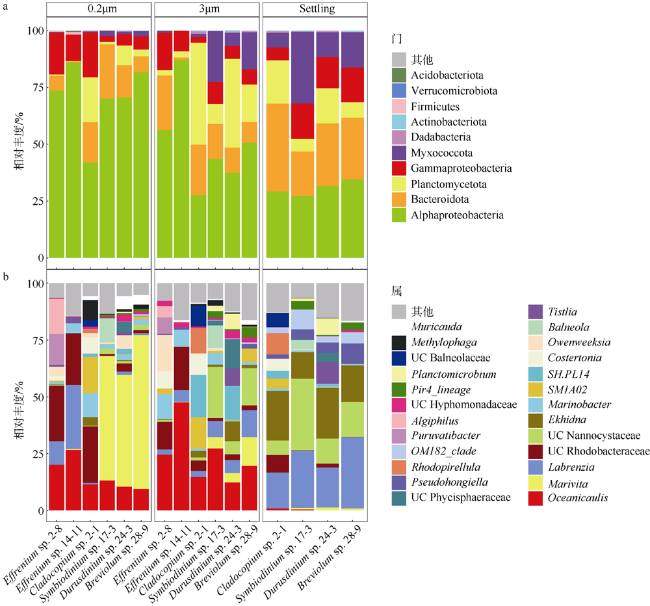
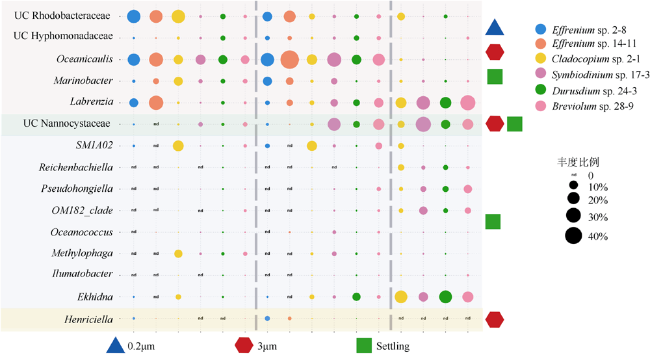
图4 虫黄藻藻际细菌群落在不同样本中的组成a. 门水平的虫黄藻藻际细菌群落物种组成, 在所有样本中总相对丰度<0.01%的属归为其他, 其中变形菌门细分为α-变形菌、β-变形菌和γ-变形菌; b. 属水平的虫黄藻藻际细菌群落组成图, 相对丰度<1%归为其他 Fig. 4 Zooxanthellae associated bacteria community composition profiles in different samples(3 μm, 0.2 μm, Settling). (a) Bacteria community composition profiles at phylum level. Relative abundance <0.01% was classified into others. Phylum Proteobacteria was subdivided into α-Proteobacteria, β-Proteobacteria and γ-Proteobacteria. (b) Bacteria community composition profiles at genus level. Relative abundance <1% was classified into others |
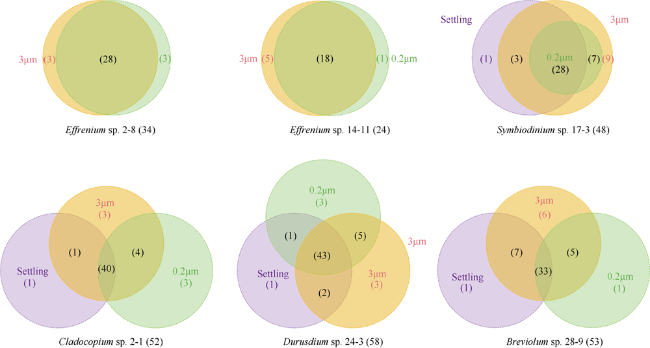
图5 虫黄藻藻际细菌基于ASV水平在不同样本(3μm、0.2μm和Settling)中的共享情况Fig. 5 Shared ASVs in different samples [3 μm, 0.2 μm and Settling (specific to anchorage-dependent living algae)] in each zooxanthellae strain |
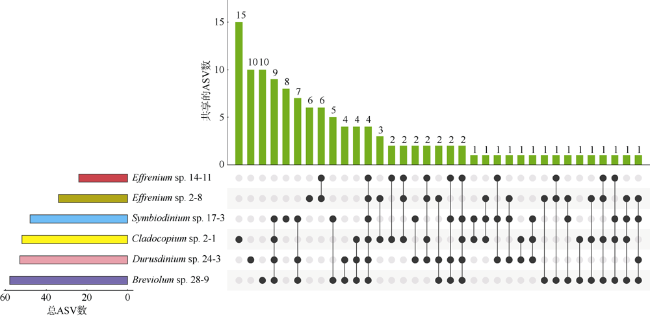
图6 不同虫黄藻细菌群落之间共享ASVs的情况Fig. 6 The UpSet diagram showing the shared ASVs between different zooxanthellae strains |
表2 不同虫黄藻株两两之间共享ASVs比例Tab. 2 The ratio of shared ASVs between different zooxanthellae strains |
| 藻株1 | 藻株2 | 两藻株共有ASVs数(取交集)/两藻株总ASVs数(取并集) | 所占比例/% |
|---|---|---|---|
| 24-3 | 28-9 | 35/76 | 46.05 |
| 17-3 | 24-3 | 33/73 | 45.21 |
| 17-3 | 28-9 | 30/71 | 42.25 |
| 2-8 | 14-11 | 16/42 | 38.10 |
| 2-1 | 28-9 | 23/82 | 28.05 |
| 2-1 | 24-3 | 24/86 | 27.91 |
| 17-3 | 2-1 | 20/80 | 25.00 |
| 14-11 | 2-1 | 14/62 | 22.58 |
| 2-8 | 24-3 | 15/71 | 21.13 |
| 14-11 | 28-9 | 11/66 | 16.67 |
| 2-8 | 24-3 | 13/79 | 16.46 |
| 14-11 | 17-3 | 10/62 | 16.13 |
| 14-11 | 24-3 | 11/71 | 15.49 |
| 2-8 | 28-9 | 10/77 | 12.99 |
| 2-8 | 17-3 | 9/73 | 12.33 |
图7 6株藻0.2μm、3μm、Settling样本核心属之间的比较四个不同颜色的区域块对应着四种类型的核心属, 自上而下依次是: 0.2μm、3μm和Settling样本的核心属; 3μm和Settling样本的核心属; Settling样本的核心属; 3μm样本的核心属 Fig. 7 Comparing the core genera in 0.2μm, 3μm, and Settling samples of 6 zooxanthellae strains. Four different colored area blocks correspond to 4 types of core genera, from top to bottom: core genera of 0.2 μm, 3 μm and Settling samples; core genera of 3μm and Settling samples; core genera of Settling samples; core genera of 3μm samples |
| [1] |
李淑, 余克服, 陈天然, 等, 2011. 珊瑚共生虫黄藻密度的季节变化及其与珊瑚白化的关系——以大亚湾石珊瑚为例[J]. 热带海洋学报, 30(2): 39-45.
|
| [2] |
吴家法, 李洁, 张偲, 2015. 鹿回头岸礁区4种造礁珊瑚中可培养细菌的多样性[J]. 广东农业科学, 42(2): 146-151.
|
| [3] |
张增虎, 唐丽丽, 张永雨, 2018. 海洋中藻菌相互关系及其生态功能[J]. 微生物学通报, 45(9): 2043-2053.
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}