
红海榄幼苗根系呼吸代谢对水淹胁迫的响应
|
程皓(1984—), 男, 安徽省岳西县人, 研究员, 从事红树林生态学研究。email: |
Copy editor: 林强
收稿日期: 2021-12-31
修回日期: 2022-04-11
网络出版日期: 2022-04-14
基金资助
国家重点研发计划项目(国家科技基础资源调查专项)(2017FY100700)
国家自然科学基金重点项目(41676086)
国家自然科学基金重点项目(U1901211)
国家自然科学基金重点项目(41876126)
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0305)
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0303)
国际伙伴计划(133244KYSB20180012)
中国科学院A类战略性先导科技专项(XDA23050200)
中国科学院A类战略性先导科技专项(XDA19060201)
Response of respiratory metabolism in the roots of Rhizophora stylosa to waterlogging
Copy editor: LIN Qiang
Received date: 2021-12-31
Revised date: 2022-04-11
Online published: 2022-04-14
Supported by
National Key Research and Development Program of China(2017FY100700)
National Natural Science Foundation of China(41676086)
National Natural Science Foundation of China(U1901211)
National Natural Science Foundation of China(41876126)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0305)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0303)
International Partnership Program of Chinese Academy of Sciences(133244KYSB20180012)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA23050200)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA19060201)
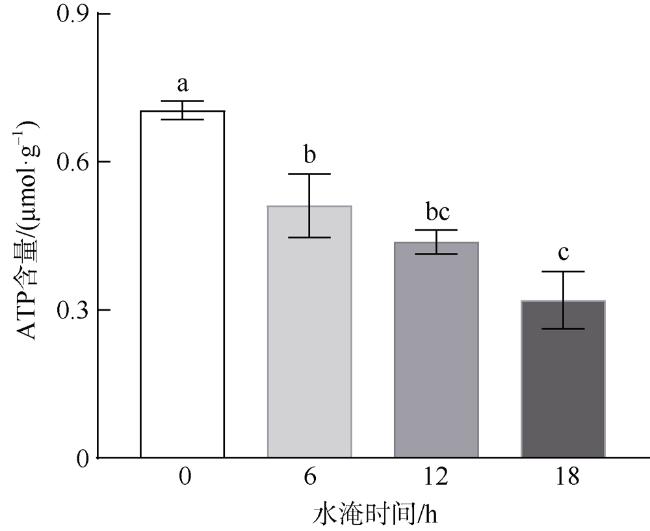
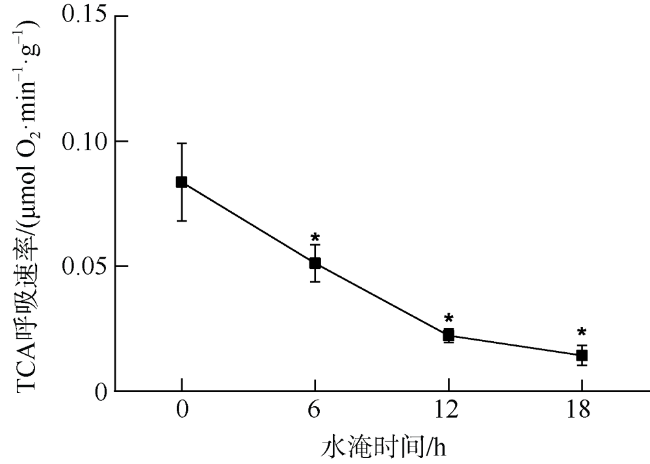
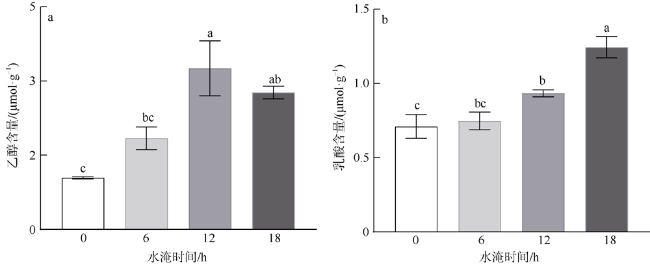
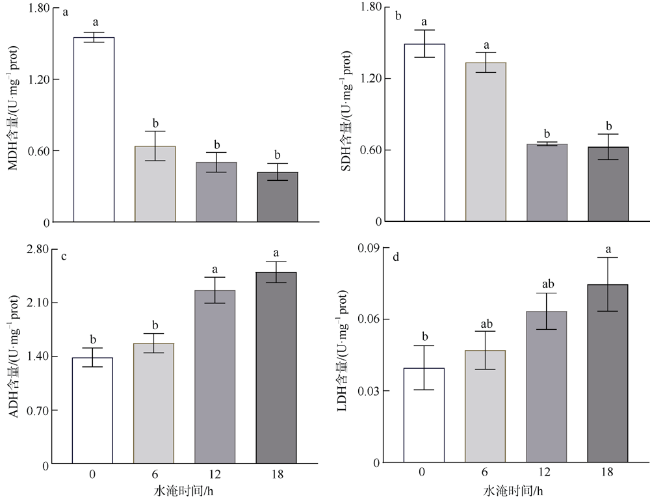
本研究利用人工潮汐系统(每天淹水0、6、12和18h), 研究了红海榄幼苗根系呼吸代谢对水淹胁迫的响应。结果表明, 每天淹水6h对红海榄幼苗的生长影响不大, 但随着水淹时间的延长, 红海榄幼苗的生长显著受到抑制。水淹时间的延长同样明显抑制了红海榄幼苗根系的产能效率, 且根系三羧酸循环的有氧呼吸速率、三磷酸腺苷含量以及琥珀酸脱氢酶和苹果酸脱氢酶活性最小值均出现在每天水淹18h处理组中。相反, 水淹胁迫下红海榄幼苗根系乙醇脱氢酶、乳酸脱氢酶活性以及乙醇和乳酸含量都呈上升的趋势。综上, 尽管无氧呼吸可一定程度上缓解三羧酸循环受阻所导致的产能匮乏, 但是过度的水淹胁迫仍会扰乱红海榄幼苗正常的生长和代谢, 导致三磷酸腺苷产能效率的降低以及无氧发酵代谢产物的积累。
程皓 , 王友绍 , 马晓宇 . 红海榄幼苗根系呼吸代谢对水淹胁迫的响应[J]. 热带海洋学报, 2022 , 41(6) : 12 -19 . DOI: 10.11978/2021189
In this study, artificial-tidal systems (0, 6, 12, 18 h·d-1) were employed to explore the effect of waterlogging on respiratory metabolism in the roots of Rhizophora stylosa. The results showed that 6 h·d-1 waterlogging treatment appeared to has little negative effect on plant growth. However, the biomass yield efficiency decreased significantly with the elongation of waterlogging duration. Tricarboxylic acid respiratory rate, adenosine-triphosphate content, and the activities of succinate dehydrogenase and malate dehydrogenase were also obviously inhibited by long-duration waterlogging, especially in the treatment of 18 h·d-1. On the contrary, alcohol dehydrogenase, lactate dehydrogenase, and the contents of alcohol and lactate increased with the elongation of waterlogging duration. In summary, although anaerobic respiration can partly alleviate energy deficiency caused by the inhibition of tricarboxylic acid cycle, excessive waterlogging would disrupt normal growth and metabolism of R. stylosa, leading to declined adenosine-triphosphate productivity and increased anaerobic fermentative metabolites.
表1 水淹胁迫下红海榄幼苗的生长以及生物量Tab. 1 Growth and biomass yield of R. stylosa subjected to waterlogging, different letters indicated significant variations between groups at P<0.05 |
| 水淹处理/(h·d-1) | 地下根系生物量/g | 地上茎叶生物量/g | 总生物量/g | 地下根系生物量/地上茎叶生物量 |
|---|---|---|---|---|
| 0 | 3.40 ± 0.18a | 4.19 ± 0.14a | 7.59 ± 0.31a | 0.81 ± 0.02a |
| 6 | 3.41 ± 0.09a | 4.45 ± 0.19a | 7.86 ± 0.26a | 0.77 ± 0.03b |
| 12 | 2.79 ± 0.14b | 4.22 ± 0.13a | 7.01 ± 0.27b | 0.66 ± 0.02c |
| 18 | 2.26 ± 0.10c | 3.79 ± 0.10b | 6.05 ± 0.20c | 0.59 ± 0.01d |
注: 不同字母代表组间存在显著性差异(P<0.05) |
| [1] |
陈思婷, 2021. 围填海对白骨壤呼吸与能量代谢的影响[J]. 广西科学院学报, 37(3): 307-313.
|
| [2] |
代捷, 程皓, 由文辉, 等, 2020. 秋茄幼苗生长生理及形态对不同潮汐处理的响应[J]. 生态科学, 39(6): 38-44.
|
| [3] |
范航清, 王文卿, 2017. 中国红树林保育的若干重要问题[J]. 厦门大学学报(自然科学版), 56(3): 323-330.
|
| [4] |
傅海峰, 陶伊佳, 王文卿, 2014. 海平面上升对中国红树林影响的几个问题[J]. 生态学杂志, 33(10): 2842-2848.
|
| [5] |
廖宝文, 邱凤英, 张留恩, 等, 2010. 红树植物白骨壤小苗对模拟潮汐淹浸时间的生长适应性[J]. 环境科学, 31(5): 1345-1351.
|
| [6] |
林鹏, 2001. 中国红树林研究进展[J]. 厦门大学学报(自然科学版), 40(2): 592-603.
|
| [7] |
刘逸泠, 覃盈盈, 郑海雷, 2017. 红树植物耐水淹和高盐适应性研究进展[J]. 厦门大学学报(自然科学版), 56(3): 314-322.
|
| [8] |
王友绍, 2019. 红树林分子生态学[M]. 北京: 科学出版社: 23-24.
|
| [9] |
王友绍, 2021. 全球气候变化对红树林生态系统的影响、挑战与机遇[J]. 热带海洋学报, 40(3): 1-14.
|
| [10] |
叶勇, 刘美龄, 卢昌义, 等, 2007. 木榄胚轴萌发及幼苗生长与生理对水渍和底质条件的响应[J]. 海洋与湖沼, 38(1): 84-90.
|
| [11] |
张艳婷, 张建军, 王建修, 等, 2016. 长期水淹对‘中山杉118’幼苗呼吸代谢的影响[J]. 植物生态学报, 40(6): 585-593.
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}