
红纤维虾形草(Phyllospadix iwatensis)种子形态及内部特征
|
韩通(1996—), 男, 内蒙古包头市人, 硕士研究生, 主要从事海草繁育与生理相关研究。email: |
Copy editor: 林强
收稿日期: 2021-11-23
修回日期: 2022-03-24
网络出版日期: 2022-04-18
基金资助
国家自然科学青年基金项目(42006110)
烟台市科技计划项目(2020MSGY058)
山东省自然科学青年基金(ZR2019QD017)
山东省重点研发计划(国际科技合作)(2019GHZ026)
联合国粮食和农业组织“中国典型河口生物多样性保护修复和保护区网络化示范项目(GCP/CPR/045/GFF)
长岛生态渔业发展与海洋牧场修复技术研究(2019MSGY126)
The seed morphology and internal characteristics of seagrass, surfgrass Phyllospadix iwatensis
Copy editor: LIN Qiang
Received date: 2021-11-23
Revised date: 2022-03-24
Online published: 2022-04-18
Supported by
National Natural Science Foundation of China(42006110)
Technology Innovation Development Program of Yantai(2020MSGY058)
Shandong Provincial Natural Science Foundation, China(ZR2019QD017)
Key R & D Projects in Shandong Province (International Scientific and Technical Cooperation)(2019GHZ026)
Food and Agriculture Organization of the United Nations “Project Management Office of Demonstration of Estuarine Biodiversity Conservation, Restoration and Protected Area Networking in China”(GCP/CPR/045/GFF)
Research on Changdao Ecological Fishery Development and Marine Ranching Restoration Technology(2019MSGY126)
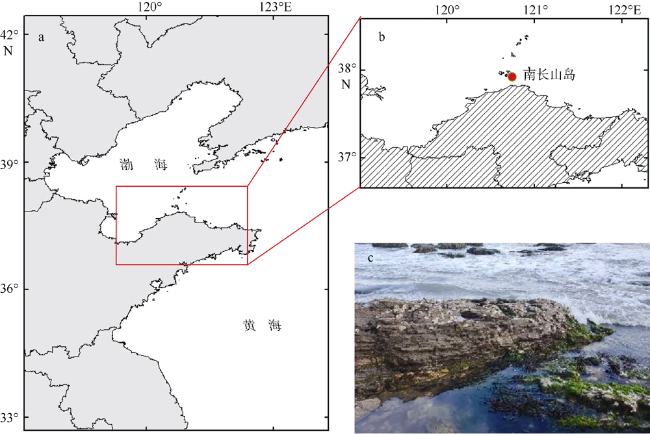
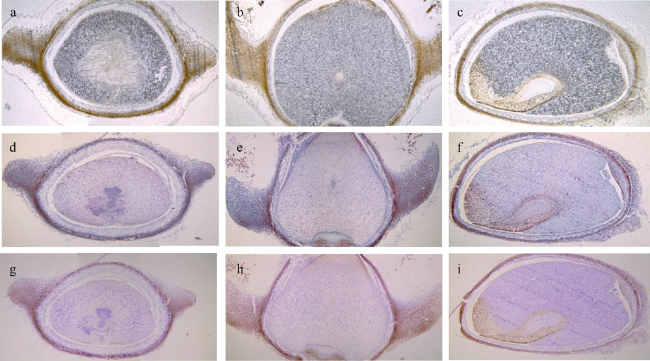
海草床可以为海洋动物提供产卵、索饵、繁育等场所。目前, 海草床面积急剧下降, 其中我国关于虾形草属(Phyllospadix)海草的研究相对较少, 对于虾形草主要繁育载体种子的描述、显微观察和萌发过程方面存在较多空白。本研究以红纤维虾形草(Phyllospadix iwatensis)为研究对象, 对其生长环境、种子外部特征、内部结构、萌发过程等进行了定性定量描述。结果表明, 虾形草果实种子对于海洋环境有着外部形态与内部结构的适应性, 外部形态特点包括独特的果序和果实形状, 以及翅状结构和内侧鬃毛, 其中果序呈弯虾形, 平均长度3.4cm, 果实呈马掌形, 直径3~4mm, 内部结构特征包括胚乳占比大、果皮种皮解剖结构复杂等, 具有作为海岸带植被修复工具种的优势。本研究补充了目前对虾形草等海洋沉水植物种子的研究资料, 为虾形草海草种质库保护与海草床修复提供了建议和参考。
韩通 , 李晶晶 , 刘正一 , 刘凯 , 张金浩 , 秦松 , 钟志海 . 红纤维虾形草(Phyllospadix iwatensis)种子形态及内部特征[J]. 热带海洋学报, 2022 , 41(6) : 105 -113 . DOI: 10.11978/2021163
The global seagrass bed ecosystems are facing multiple threats. For instance, the seagrass Phyllospadix (Zosteraceae) is of great ecological importance and fishery value. And one of the seagrasses surfgrass, Phyllospadix, is relatively poorly studied. In particular, there are more study gaps in the description, micro-observation and germination process of the main breeding carrier, seeds of surfgrass, as well as in the identification of DNA molecules. In this study, Phyllospadix iwatensis was used as the study object to describe its growing environment, external characteristics of seeds, internal structure, germination process, etc. in both qualitative and quantitative ways, and the results show that fruits and seeds of surfgrass have the adaptability on external form and internal structure in the marine environment, and have the advantage of becoming tool species for vegetation restoration in coastal zone. External morphological features include unique fruit inflorescence and fruit shape, as well as wing-like structure and medial mane, of which the fruit inflorescence is curved shrimp-shaped with an average length of 3.4 cm and the fruit is horseshoe-shaped with a diameter of 3-4 mm. The internal structural features include a large proportion of endosperm and complex anatomy of the peel and seed coat. This study complements the lack of in-depth study on the seeds of marine submerged plants, such as surfgrass, and provides valuable suggestions and reference contents for the protection of surfgrass and other types of seagrass seed banks, as well as the restoration of seagrass beds, and also yields a basis for the practical application of surfgrass in seedling restoration.
Key words: seagrass; Phyllospadix iwatensis; fruit; seed; morphology
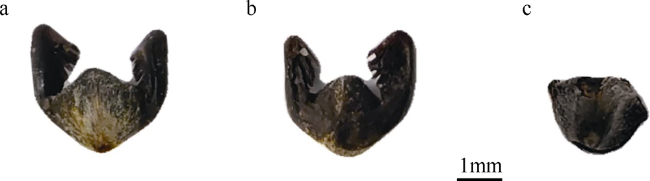
图3 红纤维虾形草(Phyllospadix iwatensis)果序形态a. 果序背面形态; b. 果序腹面形态; c. 果序未剥开外观; d. 果序剥开外观 Fig. 3 The shape of Phyllospadix iwatensis involucre |
表1 23粒红纤维虾形草果序外观描述数据统计结果Tab. 1 The statistical results of involucre shape |
| 果序长度/cm | 柄长度/cm | 果序宽度/cm | 果序厚度/cm | 种子数目 | |
|---|---|---|---|---|---|
| 平均值 | 3.4 | 2.8 | 0.6 | 0.4 | 5.6 |
| 标准差 | 0.5 | 0.6 | 0.1 | 0.1 | 0.7 |
| 标准误差 | 0.7 | 0.8 | 0.1 | 0.1 | 1.6 |
| [1] |
曹政飞, 2015. 红纤维虾海藻形态结构及其对海洋环境的适应性[D]. 济南: 山东大学.
|
| [2] |
曹政飞, 张伟, 赵宏, 2015. 红纤维虾海藻(Phyllospadix iwatensis)形态结构特征及其对海洋环境的适应性[J]. 海洋与湖沼, 46(6): 1326-1332.
|
| [3] |
陈启明, 刘松林, 张弛, 等, 2020. 海南典型热带海草床4种代表性鱼类的生长特征及其对海草资源量变化的响应[J]. 热带海洋学报, 39(5): 62-70.
|
| [4] |
房丽宁, 李青丰, 易津, 1997. 几种苔草种子的形态观察[J]. 中国草地, (5): 49-51.
|
| [5] |
黄小平, 江志坚, 范航清, 等, 2016. 中国海草的“藻”名更改[J]. 海洋与湖沼, 47(1): 290-294.
|
| [6] |
李捷, 刘译蔓, 孙辉, 等, 2019. 中国海岸带蓝碳现状分析[J]. 环境科学与技术, 42(10): 207-216.
|
| [7] |
马瑞, 李文涛, 李常军, 2019. 虾海草的研究进展[J]. 海洋湖沼通报, (2): 153-161.
|
| [8] |
潘金华, 2015. 大叶藻(Zostera marina L.)场修复技术与应用研究[D]. 青岛: 中国海洋大学.
|
| [9] |
徐少春, 2017. 山东沿海典型海草床鳗草种子生态学研究[D]. 青岛: 中国科学院大学(中国科学院海洋研究所).
|
| [10] |
杨顶田, 刘素敏, 单秀娟, 2013. 海草碳通量的卫星遥感检测研究进展[J]. 热带海洋学报, 32(6): 108-114.
|
| [11] |
杨宗岱, 1979. 中国海草植物地理学的研究[J]. 海洋湖沼通报, (2): 41-46.
|
| [12] |
于硕, 张景平, 崔黎军, 等, 2019. 基于种子法的海菖蒲海草床恢复[J]. 热带海洋学报, 38(1): 49-54.
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
/
| 〈 |
|
〉 |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}