
长棘海星(Acanthaster planci)幼体特异性PCR检测技术与应用*
|
张颖(1992—), 女, 湖南省郴州市人, 助理研究员, 从事海洋生物分子生态学研究。email: |
Copy editor: 林强
收稿日期: 2022-01-19
修回日期: 2022-05-09
网络出版日期: 2022-05-19
基金资助
财政部和农业农村部国家现代农业产业技术体系
Specific PCR detection for Acanthaster planci larvae and its application*
Copy editor: LIN Qiang
Received date: 2022-01-19
Revised date: 2022-05-09
Online published: 2022-05-19
Supported by
China Agriculture Research System of MOF and MARA
长棘海星(Acanthaster planci)的暴发是导致我国南海乃至印度—太平洋海域珊瑚礁退化的主要原因之一。浮游幼体的密度是决定成体种群是否暴发的重要指标, 但是由于幼体肉眼不可见且不易分辨, 常规调查和显微镜观察均无法有效检测到自然海域的长棘海星幼体, 因此迫切需要开发一种高灵敏性且特异性的长棘海星幼体检测技术。本研究针对长棘海星幼体线粒体细胞色素氧化酶亚基I(mtCOI, mitochondrial cytochrome oxidase subunit I)基因序列, 建立了基于聚合酶链式反应(PCR, polymerase chain reaction)的长棘海星幼体特异性检测技术, 并对西沙七连屿珊瑚礁海域的长棘海星幼体进行了检测。结果表明, 设计筛选的4对特异性引物均可以扩增长棘海星ApmtCOI基因片段, 且与蓝指海星(Linckia laevigata)、面包海星(Culcita novaeguineae)、粒皮海星(Choriaster granulatus)和吕宋棘海星(Echinaster luzonicus)没有交叉反应。在退火温度为58.5℃时, 引物2aooniF/2aooniR的特异性最佳, 具有较高的灵敏度, 可检测到皮克级的长棘海星基因组DNA。利用该技术检测了西沙七连屿海域长棘海星幼体分布情况, 发现10月底西沙七连屿珊瑚礁海域能检测到长棘海星幼体, 且幼体分布并不均匀。因此, 该检测技术可作为今后长棘海星幼体种群监测的有效方法。
关键词: 长棘海星; 细胞色素氧化酶亚基I; 聚合酶链式反应; 幼体
张颖 , 杨栎潼 , 刘冰 , 郑凡昱 , 罗鹏 , 陈偿 . 长棘海星(Acanthaster planci)幼体特异性PCR检测技术与应用*[J]. 热带海洋学报, 2022 , 41(6) : 125 -131 . DOI: 10.11978/2022011
The outbreak of crown-of-thorns seastar (CoTS, Acanthaster planci) is one of the main causes to coral reef degradation in the South China Sea and the Indo-Pacific region. The density of CoTS larvae is an important indicator to determine whether the outbreak of CoTS adult population occurs or not. However, as the larvae are not visible and thus difficult to be distinguished, conventional investigation and microscope observation cannot effectively detect the CoTS larvae in natural seawaters. Therefore, it is needed to develop a sensitive and specific method for detecting CoTS larvae. This study established specific polymerase chain reaction (PCR) method to detect the CoTS larvae based on CoTS mitochondrial cytochrome oxidase subunit I gene (ApmtCOI), and this method was applied to detect the CoTS larvae in the coral reef around Qilianyu Island, Xisha. The results showed that the designed and screened four pairs of specific primers could succeed in amplifying ApmtCOI gene, and had no cross-reaction with Linckia laevigata, Culcita novaeguineae, Choriaster granulatus and Echinaster luzonicus. Moreover, the primer 2aooniF/2anooiR had the best specificity and higher sensitivity when annealing temperature was 58.5°C, which could detect the CoTS genomic DNA in pictogram grade. Furthermore, this method had succeeded in detecting the CoTS larvae in the Qilianyu Island of Xisha at the end of October, and we found that the distribution of CoTS larvae was not uniform. Therefore, this detection technology can be used as an effective method for monitoring CoTS larvae population in the future.
表1 样品采集的站位Tab. 1 Sampling stations |
| 站位 | 坐标 | 地理位置 | 样品名 |
|---|---|---|---|
| 1 | 16°57.708′ N, 112°13.080′ E | 西沙洲 | XSZ |
| 2 | 16°58.117′ N, 112°14.848′ E | 赵述岛 | ZS-1 |
| 3 | 16°58.315′ N, 112°16.300′ E | 赵述岛 | ZS-2 |
| 4 | 16°57.112′ N, 112°19.432′ E | 中岛 | ZD |
| 5 | 16°56.794′ N, 112°19.801′ E | 南岛 | ND |
| 6 | 16°56.086′ N, 112°20.221′ E | 北沙洲 | BSZ |
表2 PCR引物Tab. 2 PCR primers |
| 序号 | 引物名称 | 引物序列(5'—3') | 熔解温度(Tm)/℃ | 线粒体基因组上的片段位点 | 目的片段 大小/bp | 来源 | |
|---|---|---|---|---|---|---|---|
| 1 | COTS-F-1321 | TCCGACTACCCGGACGCC TATAC | 62.9 | 5985—6007 | 126 | Doyle et al, 2017 | |
| COTS-R-1446 | AGTGGT TCGCTGGGAAGTGAAGG | 62.2 | 6008—6110 | ||||
| 2 | 2aooniF | ATTTGGGGCCTGAGCAGGAATG | 60.6 | 4727—4748 | 301 | Suzuki et al, 2016 | |
| 2aooniR | GCCTCTTTCTACTCCGGCTGATG | 60.2 | 5005—5027 | ||||
| 3 | AcanP-TF | GCACGATTTGTCTCTGCCAAA | 56.1 | 3074—3094 | 785 | 本研究 | |
| AcanP-TR | AGTCCTTCTCTCGCCAGGT | 58.4 | 3840—3858 | ||||
| 4 | COTS-F-69 | GGCCTGAGCAGGAATGGTTGGAA | 62.6 | 4733—4755 | 919 | Uthicke et al, 2015 | |
| COTS-R-987 | GCCTTGTAGCGTTGCCATTCACC | 61.9 | 5629—5651 | ||||
表3 PCR反应条件Tab. 3 PCR reaction conditions |
| 序号 | 引物名称 | PCR 反应条件 |
|---|---|---|
| 1 | COTS-F-1321 COTS-R-1446 | 95℃ 3min; 95℃ 15s, 60℃ 15s, 72℃ 30s, 34个循环; 72℃ 10min; |
| 2 | 2aooniF 2aooniR | 95℃ 3min; 95℃ 15s, 60℃ 15s, 72℃ 30s, 34个循环; 72℃ 10min; |
| 4 | AcanP-TF AcanP-TR | 95℃ 3min; 95℃ 15s, 57℃ 15s, 72℃ 1min, 34个循环; 72℃ 10min; |
| 5 | COTS-F-69 COTS-R-987 | 95℃ 3min; 95℃ 15s, 57℃ 15s, 72℃ 1min, 34个循环; 72℃ 10min; |
图2 不同引物的特异性检测M: 2000bp marker; 1—6分别为COST-F-1321/COST-R-1446引物扩增下的样品D5、H5、X5、L5、阴性、阳性; 7—12分别为2aooniF/2aooniR引物扩增下的样品D5、H5、X5、L5、阴性、阳性; 13—18分别为AcanP-TF/TR引物扩增下的样品D5、H5、X5、L5、阴性、阳性; 19—24分别为COTS-F-69/COTS-R-987引物扩增下的样品D5、H5、X5、L5、阴性、阳性 Fig. 2 Specificity screening of different primers. M: 2000bp marker; 1-6: D5, H5, X5, L5, negative, positive control samples were amplified by COST-F-1321/COST-R-1446 primer; 7-12: D5, H5, X5, L5, negative, positive control samples were amplified by 2aooniF/2aooniR primer; 13-18: D5, H5, X5, L5, negative, positive control samples were amplified by AcanP-TF/TR primer; 19-24: D5, H5, X5, L5, negative, positive control samples were amplified by COTS-F-69/COTS-R-987 primer |
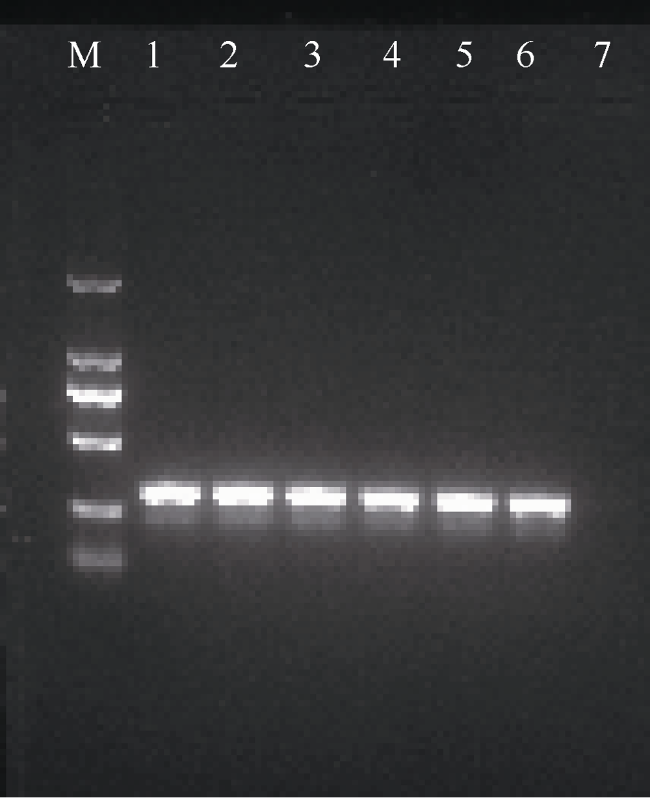
图3 2aooniF/2aooniR引物在不同退火温度下对长棘海星幼体的扩增M: 2000bp marker; 1—6分别是退火温度为56.5、57.5、58.5、59.5、60.5、61.5℃下对长棘海星幼体的扩增; 7为阴性对照 Fig. 3 Amplification of 2aooniF/2aooniR primer at different annealing temperatures on CoTS larvae. M: 2000bp marker; 1-6: the different annealing temperatures which were 56.5, 57.5, 58.5, 59.5, 60.5 and 61.5℃, respectively; 7: negative control sample |
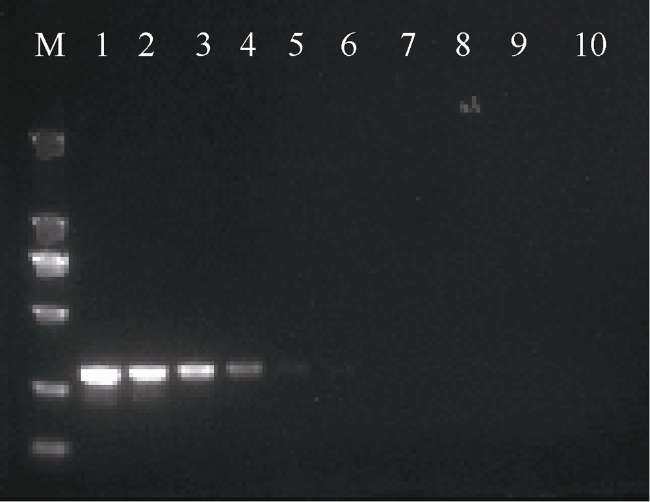
图4 2aooniF/2aooniR引物在退火温度为58.5℃下, 对不同浓度长棘海星幼体的扩增M: 2000bp marker; 1—9浓度分别为4.17、4.17×10-1、4.17×10-2、4.17×10-3、4.17×10-4、4.17×10-5、4.17×10-6、4.17×10-7、4.17×10-8ng·μL-1; 10为阴性对照 Fig. 4 Amplification of 2aooniF/2aooniR primer at 58.5℃ annealing temperatures on CoTS larvae. M: 2000bp marker; 1-9: 4.17, 4.17×10-1, 4.17×10-2, 4.17×10-3, 4.17×10-4, 4.17×10-5, 4.17×10-6, 4.17×10-7 and 4.17×10-8 ng·μL-1, respectively; 10: negative control sample |
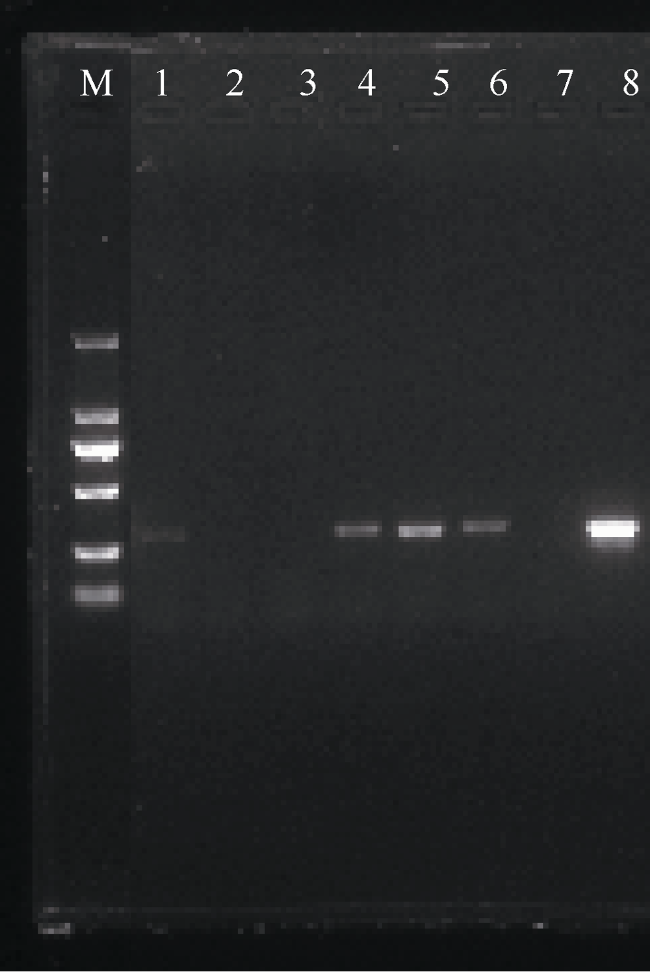
图5 2aooniF/2aooniR引物在58.5℃退火温度下对浮游生物样品的扩增M: 为2000bp marker; 1—8分别为58.5℃下扩增的XSZ、ZS-1、ZS-2、ZD、ND、BSZ、阴性、阳性样品 Fig. 5 Amplification of 2aooniF/2aooniR primer at 58.5℃ annealing temperatures on the plankton samples. M: 2000 bp marker; 1-8: XSZ, ZS-1, ZS-2, ZD, ND, BSZ, negative and positive control sample was amplified at 58.5℃ annealing temperatures, respectively |
| [1] |
李元超, 吴钟解, 梁计林, 等, 2019. 近15年西沙群岛长棘海星暴发周期及暴发原因分析[J]. 科学通报, 64(33): 3478-3484.
|
| [2] |
位正鹏, 孔晓瑜, 吴相云, 等, 2009. 基于COI序列的翡翠股贻贝Perna viridis线粒体遗传特性分析及其近缘种间的系统关系探讨[J]. 热带海洋学报, 28(6): 72-78.
|
| [3] |
吴钟解, 王道儒, 涂志刚, 等, 2011. 西沙生态监控区造礁石珊瑚退化原因分析[J]. 海洋学报, 33(4): 140-146.
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}