
基于浮游植物吸收系数和光合有效辐射的南海区域性分粒级初级生产力算法初探
|
赵红五一(1996—), 女, 硕士研究生, 从事海洋光学遥感研究。email: |
Copy editor: 姚衍桃
收稿日期: 2022-01-29
修回日期: 2022-05-29
网络出版日期: 2022-05-31
基金资助
南方海洋科学与工程广东省实验室(广州)重大专项创新团队项目(GML2019ZD0305)
南方海洋科学与工程广东省实验室(广州)重大专项创新团队项目(GML2019ZD0602)
国家自然科学基金(41976170)
国家自然科学基金(41976172)
国家自然科学基金(42276181)
国家自然科学基金(41976181)
A study of the regional size-fractionated primary production algorithm based on phytoplankton absorption coefficient and photosynthetically active radiation in the South China Sea
Copy editor: YAO Yantao
Received date: 2022-01-29
Revised date: 2022-05-29
Online published: 2022-05-31
Supported by
Major Special Innovation Team Project of Guangdong Provincial Laboratory of Southern Ocean Science and Engineering(Guangzhou)(GML2019ZD0305)
Major Special Innovation Team Project of Guangdong Provincial Laboratory of Southern Ocean Science and Engineering(Guangzhou)(GML2019ZD0602)
National Natural Science Foundation of China(41976170)
National Natural Science Foundation of China(41976172)
National Natural Science Foundation of China(42276181)
National Natural Science Foundation of China(41976181)
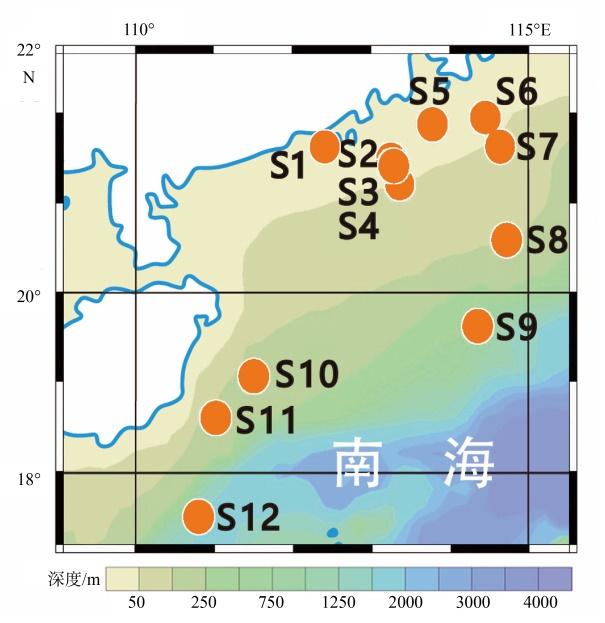
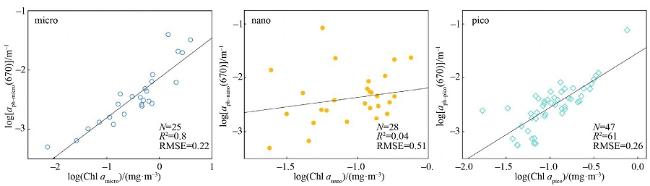
海洋初级生产过程是海洋碳循环的重要组成部分, 影响生物地球化学循环和全球气候变化。浮游植物作为海洋初级生产的主要贡献者, 按粒径大小可分为小型(micro粒级, >20μm)、微型(nano粒级, 2~20μm)和微微型(pico粒级, <2μm)。不同粒级浮游植物初级生产力(size-fractionated primary production, PPsize)对总初级生产力贡献不同, 在海洋物质能量流动及碳循环中扮演着不同角色。本文基于2019年南海西部夏季航次12个站位的生物光学剖面数据, 研究了南海西部分粒级浮游植物叶绿素a浓度和初级生产力的空间分布及它们对总叶绿素a浓度和总初级生产力的贡献百分比。利用各粒级670nm波段的浮游植物吸收系数[size-fractionated phytoplankton absorption coefficient at 670nm, aph-size(670)]与光合有效辐射(photosynthetically active radiation, PAR)的乘积[aph-size(670)×PAR]建立了南海分粒级初级生产力算法, 对于小型、微型和微微型浮游植物数据集, log[aph-size(670)×PAR]与log(PPsize)之间的决定系数R2分别为0.64、0.76和0.67。交叉验证的结果表明, 该算法具有良好的泛化性能。其性能显著优于仅利用浮游植物吸收系数估算分粒级初级生产力的算法, 表明PAR是影响分粒级初级生产力变化的重要因素之一。采用基于叶绿素a浓度的算法估算各粒级初级生产力时, 针对小型和微微型浮游植物数据集, 该算法的性能与本文构建的算法近似; 但针对微型浮游植物数据集时, 基于叶绿素a浓度的算法性能显著较低, 这可能归因于微型浮游植物吸收系数与叶绿素a浓度间的弱相关性。
关键词: 分粒级浮游植物吸收系数; 分粒级浮游植物初级生产力; 光合有效辐射; 分粒级叶绿素a
赵红五一 , 周雯 , 曾凯 , 邓霖 , 廖健祖 , 曹文熙 . 基于浮游植物吸收系数和光合有效辐射的南海区域性分粒级初级生产力算法初探[J]. 热带海洋学报, 2023 , 42(1) : 43 -55 . DOI: 10.11978/2022019
Marine primary production is an important part of the ocean carbon cycle, affecting biogeochemical cycles and global climate change. Phytoplankton, as the main contributor to marine primary production, can be classified as micro- (>20μm), nano- (2~20μm), and pico- (<2μm) phytoplankton depending on particle size. Different phytoplankton size classes contribute differently to primary production (PPsize) and thus play different roles in the oceanic circulation of matter or energy and ocean carbon cycle. Based on the bio-optical dataset collected at 12 stations in the western South China Sea in 2019, this study presented the spatial variability of size-fractionated primary production and chlorophyll a concentration of phytoplankton and their percentage contribution. The size-fractionated primary productivity of phytoplankton was well estimated from the product of size-fractionated phytoplankton absorption coefficient at 670nm [aph-size(670)] and photosynthetically active radiation (PAR) [aph-size(670)×PAR]. The coefficients of determination R2 between log[aph-size(670)×PAR] and log(PPsize) were 0.64, 0.76, and 0.67 for the micro-, nano-, and pico-phytoplankton dataset, respectively. The cross-validation of the algorithm based on the size-fractionated phytoplankton absorption coefficient and PAR has shown a good generalization performance. This algorithm could better predict the size-fractionated primary productivity compared to the size-fractionated phytoplankton absorption coefficient as the only input. This result indicates that PAR is one of the important factors to estimate the size-fractionated primary productivity. Meanwhile, the performance of the chlorophyll a concentration-based algorithm for estimating primary productivity at each size was closer to that of the algorithm constructed in this paper for both micro- and pico- phytoplankton dataset, but significantly lower for the nano-phytoplankton, probably due to the weak correlation between the absorption coefficients and chlorophyll a concentration of nano-phytoplankton.
表1 三类初级生产力算法的基本公式及输入参量Tab. 1 Formulas and input parameters of three types of primary productivity algorithms |
| 初级生产力算法类型 | 公式 | 参量意义 |
|---|---|---|
| 基于浮游植物叶绿素a浓度 | PP=Chl a×Q×R/K | PP为浮游植物初级生产力; Chl a为浮游植物叶绿素a浓度; Q为在光饱和情况下, 单位时间内单位质量叶绿素a同化的碳量; R是相对光合速率, 随海表光照强度变化; K是每米水层消光系数 |
| 基于浮游植物吸收系数 | PP=ϕ×aph×E | PP为浮游植物初级生产力; aph为浮游植物吸收系数; ϕ为光合作用光量子产率; E为光照强度 |
| 基于浮游植物碳 | PP=C×μ×Zeu×f(I0) | PP为浮游植物初级生产力; C为浮游植物碳; μ为生长速率; Zeu为真光层深度; f(I0)是表面光强的函数, 描述了固碳速率的剖面分布 |
表2 各粒级浮游植物群落对近岸珠江口海域及外海海域各水层的初级生产力和叶绿素a浓度的贡献Tab. 2 Each water layer contributions of each size phytoplankton communities to the primary productivity and chlorophyll a concentration in the nearshore Pearl River Estuary and offshore waters |
| PAR衰减百分比 | 近岸珠江口海域 | 外海海域 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| micro占比/% | nano占比/% | pico占比/% | micro占比/% | nano占比/% | pico占比/% | ||||||||
| 均值±$\sigma$ | 范围 | 均值±$\sigma$ | 范围 | 均值±$\sigma$ | 范围 | 均值±$\sigma$ | 范围 | 均值±$\sigma$ | 范围 | 均值±$\sigma$ | 范围 | ||
| 初级生产力 | 0% | 36±2 | 19~51 | 31±0.4 | 20~42 | 33±1 | 18~52 | 6±0.05 | 2~8 | 28±0.6 | 23~41 | 67±0.7 | 54~74 |
| 44% | 21±0.4 | 15~28 | 30±2 | 16~37 | 49±3 | 35~69 | 3±0.006 | 2~4 | 20±0.2 | 14~28 | 77±0.3 | 68~83 | |
| 78% | 25±3 | 3~38 | 25±1 | 11~39 | 50±5 | 22~86 | 3±0.01 | 1~5 | 15±0.1 | 13~21 | 82±0.1 | 76~84 | |
| 93% | 16±2 | 2~36 | 20±1 | 9~33 | 63±6 | 31~89 | 18±2 | 2~41 | 22±2 | 8~40 | 60±9 | 19~90 | |
| 99% | 21±1 | 9~42 | 30±1 | 19~48 | 48±1 | 38~69 | 18±1 | 9~38 | 27±2 | 15~50 | 55±5 | 29~75 | |
| 叶绿素a | 0% | 74±1 | 58~88 | 11±0.1 | 7~16 | 16±0.8 | 5~28 | 11±0.03 | 8~13 | 23±0.2 | 18~28 | 66±0.4 | 60~73 |
| 44% | 57±2 | 50~71 | 16±0.2 | 12~20 | 26±0.7 | 17~33 | 10±0.008 | 9~11 | 23±0.06 | 20~26 | 67±0.1 | 63~71 | |
| 78% | 55±1 | 17~84 | 18±0.9 | 7~30 | 27±3 | 9~54 | 11±0.02 | 9~12 | 24±0.03 | 23~26 | 65±0.1 | 62~69 | |
| 93% | 53±10 | 13~92 | 21±2 | 1~42 | 26±3 | 7~45 | 32±4 | 17~64 | 31±0.2 | 22~34 | 37±3 | 14~50 | |
| 99% | 57±7 | 28~91 | 19±2 | 2~34 | 24±3 | 6~47 | 39±3 | 19~61 | 29±0.2 | 23~34 | 32±2 | 16~50 | |
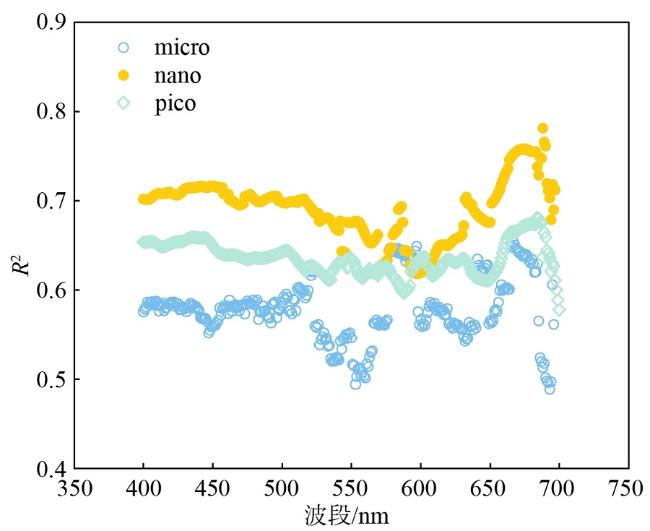
图2 各粒级浮游植物群落log[aph(λ0)×PAR]与log(PP)的线性拟合决定系数(R2)随波长的变化Fig. 2 R2 of each particle size phytoplankton communities for each band between log[aph(λ0)×PAR] and log(PP) |
表3 各粒级浮游植物吸收波段的log[aph-size(λ0)×PAR]与log(PPsize)之间的线性回归参数和评价指标Tab. 3 Linear regression parameters and evaluation indicators for the linear fitting of each particle size between log[aph-size(λ0)×PAR] and log(PPsize) |
| 参与建模的波段 | 回归参数和评价指标 | micro | nano | pico |
|---|---|---|---|---|
| 443nm | $\quad\textit{R}^2_{443}$ | 0.57 | 0.72 | 0.66 |
| k443 | 0.69 | 0.56 | 0.59 | |
| b443 | -2.8 | -3.1 | -3.0 | |
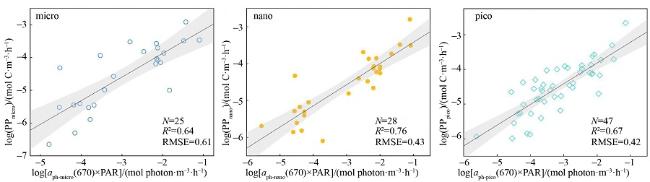
| 670nm | $\quad\textit{R}^2_{670}$ | 0.64 | 0.76 | 0.67 |
| k670 | 0.72 | 0.57 | 0.60 | |
| b670 | -2.4 | -2.8 | -2.6 | |
| 线性拟合最高R2及对应的k、b | $\quad\textit{R}^2_{max}$ | 0.65(667nm) | 0.78(688nm) | 0.68(684nm) |
| kmax | 0.73(667nm) | 0.59(688nm) | 0.58(684nm) | |
| bmax | -2.4(667nm) | -2.6(688nm) | -2.6(684nm) |
注: R2、k、b分别表示对应的决定系数、斜率和截距; 、kmax、bmax后括注的数据为线性拟合最高R2所在的波段 |
图3 各粒级浮游植物吸收波段log[aph-size(λ0)×PAR]与log(PPsize)的线性拟合图黑实线为拟合线, 灰色条带表示95%置信区间 Fig. 3 Linear fit of log[aph-size(λ0)×PAR] to log(PPsize) for phytoplankton absorption bands at each grain level. The solid black line is the fitted line, and the grey bars represent 95% confidence intervals |
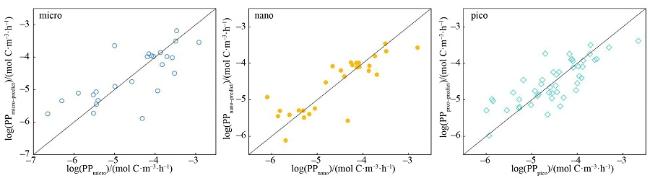
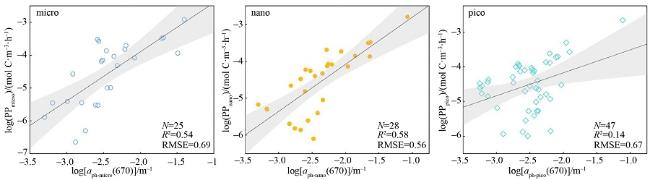
图 5 仅利用浮游植物吸收系数估算的各粒级初级生产力的分布黑实线为拟合线, 灰色条带表示95%置信区间 Fig. 5 The performance of primary productivity was predicted just using phytoplankton absorption coefficients at each size fractionated phytoplankton. The solid black line is the fitted line, and the grey bars represent 95% confidence intervals |
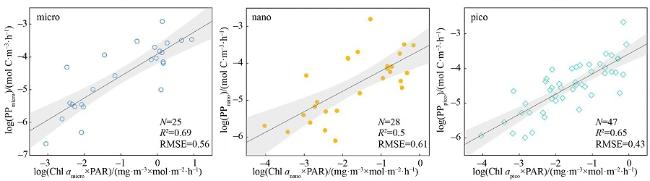
图6 利用叶绿素a和光合有效辐射(PAR)估算的各粒级初级生产力的分布黑实线为拟合线, 灰色条带表示95%置信区间 Fig. 6 The performance of primary productivity was predicted using Chl a and PAR at each size fractionated phytoplankton. The solid black line is the fitted line, and the grey bars represent 95% confidence intervals |
表 4 三种分粒级初级生产力算法的评价指标对比Tab. 4 Comparison of three size-fractionated primary productivity algorithms |
| 算法 | micro | nano | pico | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| R2 | RMSE | N | R2 | RMSE | N | R2 | RMSE | N | ||
| $\log(\mathrm{PP})=k\log\left[a_{\mathrm{bh}}(\lambda_0)\times\mathrm{PAR}\right]+b$ | 0.64 | 0.61 | 25 | 0.76 | 0.43 | 28 | 0.67 | 0.42 | 47 | |
| $\log(\mathrm{PP})=k\log\left[a_{\mathrm{bh}}(\lambda_0)\times\mathrm{PAR}\right]+b$ | 0.54 | 0.69 | 25 | 0.58 | 0.56 | 28 | 0.14 | 0.67 | 47 | |
| $\log(\mathrm{PP}_2)=k_2\log(\mathrm{Chl}a\times\mathrm{PAR})+b_2$ | 0.69 | 0.56 | 25 | 0.50 | 0.61 | 28 | 0.65 | 0.43 | 47 | |
注: 为本研究所搭建的算法, 为仅利用浮游植物吸收系数搭建的分粒级初级生产力算法, 为使用叶绿素a估算分粒级初级生产力的算法。k、k1、k2为3个线性算法的线性拟合斜率, b、b1、b2为对应的截距 |
| [1] |
傅明珠, 王宗灵, 李艳, 等, 2009. 胶州湾浮游植物初级生产力粒级结构及固碳能力研究[J]. 海洋科学进展, 27(3): 357-366.
|
| [2] |
龚祝清, 2020. 北部湾近岸海洋初级生产力遥感估算研究[D]. 南宁: 广西大学.
|
| [3] |
黄邦钦, 洪华生, 柯林, 等, 2005. 珠江口分粒级叶绿素a和初级生产力研究[J]. 海洋学报, 27(6): 180-186.
|
| [4] |
金松, 韩震, 李雪娜, 等, 2017. 叶绿素浓度和海表温度与黄海绿潮海洋初级生产力关系的研究[J]. 海洋湖沼通报, 39(3): 131-138.
|
| [5] |
孙强, 洪娅岚, 鹿凤, 2019. 珠江口邻域海洋初级生产力与人类活动相关性研究[J]. 科技创新与生产力, (3): 12-15.
|
| [6] |
唐世林, 陈楚群, 詹海刚, 2006. 海洋初级生产力的遥感研究进展[J]. 台湾海峡, 25(4): 591-598.
|
| [7] |
王荣, 焦念志, 李超伦, 等, 1995. 胶州湾的初级生产力和新生产力[J]. 海洋科学集刊, 995(1): 181-194.
|
| [8] |
薛冰, 孙军, 李婷婷, 2016. 2014年夏季南海北部浮游植物群落结构[J]. 海洋学报, 38(4): 54-65.
|
| [9] |
杨威, 董园, 俎婷婷, 等, 2019. 南海北部夏季叶绿素a分布规律及影响因素[J]. 热带海洋学报, 38(6): 9-20.
|
| [10] |
翟红昌, 2010. 基于HPLC技术的南海北部浮游植物色素与群落结构研究[D]. 青岛: 中国海洋大学.
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}