
海参产卵行为的调控激素研究进展
|
罗鹏(1977—), 男, 湖北省十堰市人, 研究员, 博士, 研究方向为海洋经济动物增养殖。email: |
Copy editor: 殷波
收稿日期: 2022-04-08
修回日期: 2022-05-25
网络出版日期: 2022-06-08
基金资助
国家重点研发计划“蓝色粮仓科技创新”专项(2020YFD0901104)
Research progress of the regulating hormones for spawning behaviors of sea cucumbers
Copy editor: YIN Bo
Received date: 2022-04-08
Revised date: 2022-05-25
Online published: 2022-06-08
Supported by
National Key Research and Development Program “Scientific and technological innovation in blue granary” Special Project(2020YFD0901104)
罗鹏 , 尹佳悦 , 马波 , 吕颖 , 任春华 , 胡超群 , 江晓 . 海参产卵行为的调控激素研究进展[J]. 热带海洋学报, 2023 , 42(2) : 178 -183 . DOI: 10.11978/2022070
Sea Cucumbers play an important role in marine ecological system and have edible and medicinal values. At present, wild resources of sea cucumbers are greatly depleted in China, and artificial breeding of sea cucumbers provides an essential way to restore the population resources. Understanding the hormones that regulate spawning behaviors of sea cucumbers has great significance for the development of artificial injection hormone. We reviewed the hormones found in sea cucumbers that can regulate their spawning behaviors, and we also provide some new perceptions for the future research. The review can provide some valuable references for revealing the key spawning mechanism of sea cucumbers and developing the artificial spawning induction by injecting hormones.
Key words: sea cucumbers; spawning; hormones; regulation
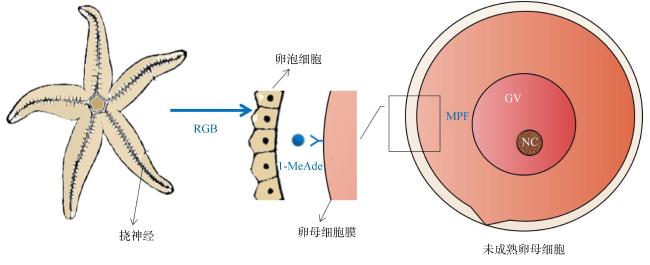
图1 海星产卵的激素调控关键机制示意图RGP: 松弛素样性腺刺激肽; 1-MeAde: 1-甲基腺嘌呤; MPF: 成熟促进因子; GV: 生长囊泡; NC: 细胞核 Fig.1 Schematic of the key mechanism of hormone regulation of starfish spawning. RGP: relaxin-like gonad-stimulating peptide; 1-MeAde: 1-methyladenine; MPF: maturation promoting factor; GV: Germinal vesicle; NC: cell nucleus |
| [1] |
陈廷, 任春华, 吴菲菲, 等, 2019. 小疣刺参促排卵短肽及其编码基因与应用[P]. 中国专利: ZL 201911120836.4.
|
| [2] |
宋坚, 庞振国, 常亚青, 等, 2009. 体内诱导刺参产卵的方法[P]. 中国专利: ZL 200910012325.0.
|
| [3] |
张春云, 王印庚, 荣小军, 等, 2004. 国内外海参自然资源、养殖状况及存在问题[J]. 海洋水产研究, 25(3): 89-97.
|
| [4] |
邹峥嵘, 易杨华, 张淑瑜, 等, 2004. 海参皂苷研究进展[J]. 中国海洋药物, 23(1): 46-53.
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}