
铵态氮加富对贝克喜盐草光合作用、谷氨酰胺合成酶和氨基酸成分的影响
|
江志坚(1982—), 男, 广东省清远市人, 研究员, 从事海草生物生态研究。email: |
Copy editor: 林强
收稿日期: 2022-05-26
修回日期: 2022-07-16
网络出版日期: 2022-07-21
基金资助
海南省科技专项(ZDYF2021SHFZ254)
国家自然科学基金项目(41735029)
国家自然科学基金项目(41976144)
国家自然科学基金项目(U1901221)
国家自然科学基金项目(42176158)
中国科学院青年创新促进会(2023359)
广东省应用海洋生物学重点实验室运行经费(2020B1212060058)
Effects of ammonium enrichment on the photosynthesis, glutamine synthetase and amino acid composition of seagrass Halophila beccarii Asch
Copy editor: LIN Qiang
Received date: 2022-05-26
Revised date: 2022-07-16
Online published: 2022-07-21
Supported by
Hainan Province Science and Technology Special Fund(ZDYF2021SHFZ254)
National Natural Science Foundation of China(41735029)
National Natural Science Foundation of China(41976144)
National Natural Science Foundation of China(U1901221)
National Natural Science Foundation of China(42176158)
Youth Innovation Promotion Association CAS(2023359)
Science and Technology Planning Project of Guangdong Province, China(2020B1212060058)
海草贝克喜盐草(Halophila beccarii)由于体型小, 其重要性一直被忽视, 且近海氮负荷增加导致其处于加速退化状态。目前贝克喜盐草对铵毒害的生理响应尚不清楚。基于室内模拟实验, 设置了四种铵态氮梯度(对照; 25、50和100μmol·L-1), 结合叶绿素荧光技术、非损伤微测技术和靶向代谢组学, 探讨了铵态氮加富对贝克喜盐草光合作用、叶绿素、叶肉细胞铵离子流速、谷氨酰胺合成酶活性以及营养成分的影响。结果表明, 贝克喜盐草叶片的最大相对电子传递速率呈现低铵态氮加富>中铵态氮加富>对照>高铵态氮加富的变化趋势, 高铵态氮加富显著降低了最大相对电子传递速率和光能利用效率, 进而减少碳库用于铵态氮的同化。同时, 铵态氮加富显著增加了铵离子内流流速和谷氨酰胺合成酶活性, 把过多的铵同化成氨基酸。但是, 铵态氮加富却降低了氨基酸成分, 这可能是由于氨基酸被用来合成有机物如关键次生代谢物, 以进一步调节和适应铵毒害作用。因此, 适度的铵营养盐增加可促进贝克喜盐草的光合作用和生长, 而高浓度的铵营养盐则对贝克喜盐草产生毒害作用。
江志坚 , Chanaka Isuranga PREMARATHNE , 方扬 , 林基桢 , 吴云超 , 刘松林 , 黄小平 . 铵态氮加富对贝克喜盐草光合作用、谷氨酰胺合成酶和氨基酸成分的影响[J]. 热带海洋学报, 2023 , 42(3) : 116 -125 . DOI: 10.11978/2022121
Seagrass Halophila beccarii Asch has long been neglected due to its small size, and the coastal nitrogen loading accelerates its degradation. The physiological response of H. beccarii to ammonium toxicity is not clear. Based on the lab cultivation experiment, the effects of ammonium enrichment in four gradients (Control, 25 μmol·L-1, 50 μmol·L-1 and 100 μmol·L-1) on the photosynthesis, chlorophyll, ammonium ion flux in mesophyll cell, glutamine synthetase activity and nutrients of H. beccarii are studied using chlorophyll fluorescence, non-invasive micro measurement technology and targeted metabolomics. The results showed that the maximum relative electron transport rate exhibits a trend of low ammonium enrichment>moderate ammonium enrichment>control>high ammonium enrichment. High ammonium enrichment significantly reduces the maximum relative electron transport rate and light utilization efficiency, while decreasing the carbon pool for ammonium assimilation. Meanwhile, ammonium enrichment significantly enhances the ammonium ion flux and glutamine synthetase activity, assimilating excess ammonium into amino acids. However, ammonium enrichment reduces the amino acid contents, which might be caused by that the amino acids are used to synthesize organic substances such as key secondary metabolites to further regulate and adapt to ammonium toxicity. Therefore, moderate ammonium enrichment is beneficial for the photosynthesis and growth of H. beccarii, while high ammonium enrichment has toxic effect on H. beccarii.
Key words: ammonium; Halophila beccarii Asch; photosynthesis; glutamine synthetase; amino acid
Chl a (μg·mL-1) = 12.7OD663 - 2.69OD645
Chl b (μg·mL-1) = 22.9OD645 - 4.86OD663
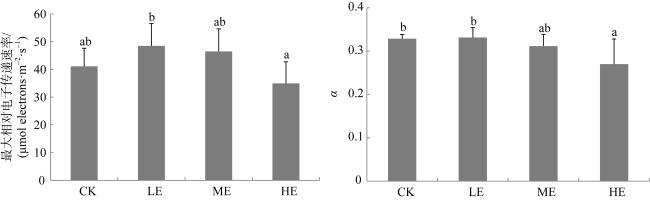
图1 铵态氮加富对贝克喜盐草最大相对电子传递速率和α(光能利用效率)的影响CK代表对照, LE代表低铵态氮加富, ME代表中铵态氮加富, HE代表高铵态氮加富。图中不同的小写字母表示差异显著(P<0.05) Fig. 1 The effect of ammonium enrichment on the relative maximum electron transport rates of Halophila beccarii. CK represents the control, LE represents low ammonium enrichment (25 μmol·L-1 NH+ 4), ME represents medium ammonium enrichment (50 μmol·L-1 NH+ 4), HE represents high ammonium enrichment (100 μmol·L-1 NH+ 4). Different small letters in the columns indicate significant differences (P<0.05) |
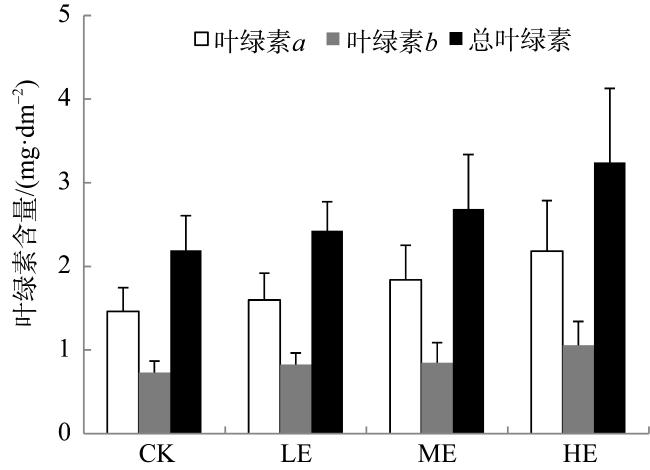
图2 铵态氮加富对贝克喜盐草叶绿素含量的影响CK代表对照, LE代表低铵态氮加富, ME代表中铵态氮加富, HE代表高铵态氮加富 Fig. 2 The effect of ammonium enrichment on the chlorophyll contents of Halophila beccarii. CK represents the control, LE represents low ammonium enrichment (25 μmol·L-1 NH+ 4), ME represents medium ammonium enrichment (50 μmol·L-1 NH+ 4), HE represents high ammonium enrichment (100 μmol·L-1 NH+ 4) |
表1 铵态氮加富对贝克喜盐草叶片氨基酸成分的影响Tab. 1 Effect of ammonium enrichment on the amino acid compositions in the leaf of Halophila beccarii |
| 氨基酸 | 对照 | 25μmol·L-1 NH+ 4 | 50μmol·L-1 NH+ 4 | 100μmol·L-1 NH+ 4 | ||||
|---|---|---|---|---|---|---|---|---|
| 含量/(μg·g-1) | 占比/% | 含量/(μg·g-1) | 占比/% | 含量/(μg·g-1) | 占比/% | 含量/(μg·g-1) | 占比/% | |
| 甘氨酸 | ND | ND | ND | ND | ND | ND | ND | ND |
| 丙氨酸 | 2101.7±710.2A | 10.35 | 1538.0±524.9AB | 9.88 | 1537.1±412.8AB | 9.54 | 1062.9±272.8B | 9.48 |
| γ-氨基丁酸 | 148.5±85.5A | 0.73 | 92.8±34.9AB | 0.60 | 100.2±53.1AB | 0.62 | 61.1±42.2B | 0.54 |
| 丝氨酸 | 254.7±70.4 | 1.25 | 218.1±62.1 | 1.40 | 218.9±55.3 | 1.36 | 154.8±46.2 | 1.38 |
| 脯氨酸 | 44.4±12.4 | 0.22 | 37.5±20.4 | 0.24 | 36.0±7.5 | 0.22 | 25.9±13.7 | 0.23 |
| 缬氨酸 | 299.0±93.9 | 1.47 | 203.2±49.6 | 1.31 | 224.7±107.4 | 1.39 | 154.8±72.7 | 1.38 |
| 苏氨酸 | ND | ND | ND | ND | ND | ND | ND | ND |
| 异亮氨酸 | 347.7±112.2 | 1.71 | 271.5±93.0 | 1.74 | 293.7±134.6 | 1.82 | 204.6±85.0 | 1.82 |
| 亮氨酸 | 75.1±20.8 | 0.37 | 49.2±9.5 | 0.32 | 61.4±40.3 | 0.38 | 36.3±19.7 | 0.32 |
| 天冬酰胺 | 14022.5±3810.7 | 69.04 | 11103.8±2500.6 | 71.34 | 11339.8±5312.9 | 70.38 | 7923.3±4034.9 | 70.64 |
| 鸟氨酸 | 2.3±1.0 | 0.01 | 3.3±1.7 | 0.02 | 2.8±1.5 | 0.02 | 1.8±0.9 | 0.02 |
| 天冬氨酸 | 506.9±114.1A | 2.50 | 392.8±65.9AB | 2.52 | 370.1±147.3AB | 2.30 | 285.4±124.3B | 2.54 |
| 高半胱氨酸 | ND | ND | ND | ND | ND | ND | ND | ND |
| 谷氨酰胺 | 2233.8±787.1A | 11.00 | 1429.1±570.5A | 9.18 | 1685.2±970.4AB | 10.46 | 1114.8±671.0B | 9.94 |
| 赖氨酸 | 11.1±2.4 | 0.05 | 14.6±8.9 | 0.09 | 10.2±6.3 | 0.06 | 6.4±4.5 | 0.06 |
| 谷氨酸 | 156.4±12.5 | 0.77 | 133.9±18.2 | 0.86 | 154.2±44.0 | 0.96 | 124.1±44.6 | 1.11 |
| 甲硫氨酸 | ND | ND | ND | ND | ND | ND | ND | ND |
| 组氨酸 | 16.9±3.0 | 0.08 | 13.3±2.8 | 0.09 | 12.4±9.8 | 0.08 | 11.0±6.7 | 0.10 |
| 苯丙氨酸 | 46.5±11.9A | 0.23 | 38.6±4.8AB | 0.25 | 39.1±19.3AB | 0.24 | 28.7±11.0B | 0.26 |
| 精氨酸 | 1.9±0.4 | 0.01 | 2.4±0.9 | 0.02 | 2.0±0.8 | 0.01 | 2.0±1.1 | 0.02 |
| 酪氨酸 | ND | ND | ND | ND | ND | ND | ND | ND |
| 色氨酸 | 41.6±14.9A | 0.20 | 23.1±7.7AB | 0.15 | 24.5±19.1B | 0.15 | 18.1±12.9B | 0.16 |
| 总氨基酸 | 20311.0±5768.9 | 100.00 | 15565.3±3627.4 | 100.00 | 16112.4±7267.6 | 100.00 | 11216.1±5384.5 | 100.00 |
注: 上标不同的大写字母表示差异显著(P<0.05) |
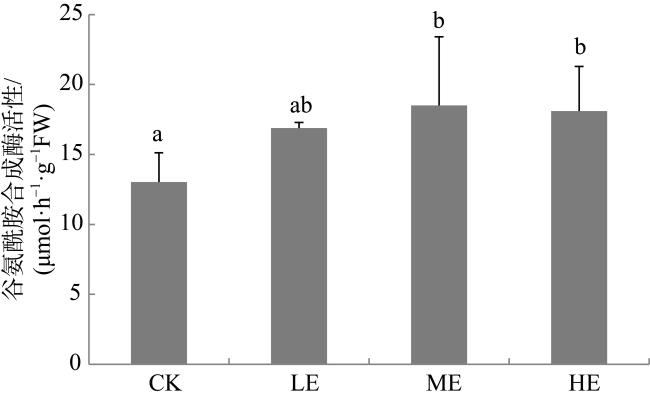
图3 铵态氮加富对贝克喜盐草叶片谷氨酰胺合成酶活性的影响CK代表对照, LE代表低铵态氮加富, ME代表中铵态氮加富, HE代表高铵态氮加富。图中不同的小写字母表示差异显著(P<0.05) Fig. 3 The effect of ammonium enrichment on the glutamine synthetase activity of the leaf of Halophila beccarii. CK represents the control, LE represents low ammonium enrichment (25 μmol·L-1 NH+ 4), ME represents medium ammonium enrichment (50 μmol·L-1 NH+ 4), HE represents high ammonium enrichment (100 μmol·L-1 NH+ 4). Different small letters in the columns indicate significant differences (P<0.05) |
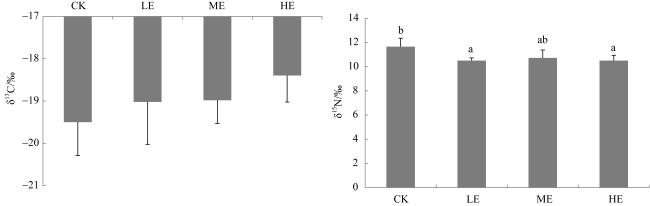
图4 铵态氮加富对贝克喜盐草叶片稳定碳、氮同位素的影响CK代表对照, LE代表低铵态氮加富, ME代表中铵态氮加富, HE代表高铵态氮加富。图中不同的小写字母表示差异显著(P<0.05) Fig. 4 The effect of ammonium enrichment on the stable isotope carbon and nitrogen of the leaf of Halophila beccarii. CK represents the control, LE represents low ammonium enrichment (25 μmol·L-1 NH+ 4), ME represents medium ammonium enrichment (50 μmol·L-1 NH+ 4), HE represents high ammonium enrichment (100 μmol·L-1 NH+ 4). Different small letters in the columns indicate significant differences (P<0.05) |
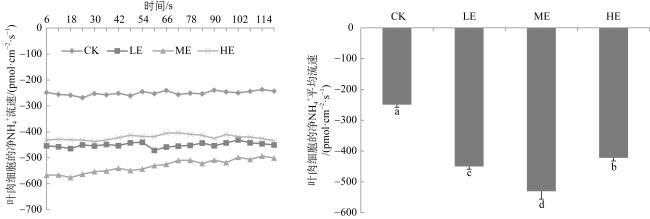
图5 铵态氮加富对贝克喜盐草叶片铵根离子流速的影响CK代表对照, LE代表低铵态氮加富, ME代表中铵态氮加富, HE代表高铵态氮加富。图中不同的小写字母表示差异显著(P<0.05) Fig. 5 The effect of ammonium enrichment on the ammonium ion flux of the leaf of Halophila beccarii. CK represents the control, LE represents low ammonium enrichment (25 μmol·L-1 NH+ 4), ME represents medium ammonium enrichment (50 μmol·L-1 NH+ 4), HE represents high ammonium enrichment (100 μmol·L-1 NH+ 4). Different small letters in the columns indicate significant differences (P<0.05) |
| [1] |
陈启明, 刘松林, 张弛, 等, 2020. 海南典型热带海草床4种代表性鱼类的生长特征及其对海草资源量变化的响应[J]. 热带海洋学报, 39(5): 62-70.
|
| [2] |
黄小平, 江志坚, 张景平, 等, 2018. 全球海草的中文命名[J]. 海洋学报, 40(4): 127-133.
|
| [3] |
李玲兰, 江志坚, 方扬, 等, 2020. 海草组学研究进展[J]. 科学通报, 65: 4063-4072.
|
| [4] |
邱广龙, 苏治南, 范航清, 等, 2020. 贝克喜盐草的生物学和生态学特征及其保护对策[J]. 海洋环境科学, 39(1): 121-126.
|
| [5] |
邱广龙, 苏治南, 钟才荣, 等, 2016. 濒危海草贝克喜盐草在海南东寨港的分布及其群落基本特征[J]. 广西植物, 36(7): 882-889.
|
| [6] |
舒展, 张晓素, 陈娟, 等, 2010. 叶绿素含量测定的简化[J]. 植物生理学通讯, 46(4): 399-402.
|
| [7] |
王道儒, 吴钟解, 陈春华, 等, 2012. 海南岛海草资源分布现状及存在威胁[J]. 海洋环境科学, 31(1): 34-38.
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}