
粤港澳大湾区近海中小型浮游动物分布特征及影响因素*
|
宋星宇(1978—), 男, 研究员, 江苏省泗洪县人, 从事海洋生态学研究。email: |
|
*本研究得到中国科学院岛礁综合研究中心的支持。感谢黄彬彬、苏芯莹、符小明、崔海平、张子韬等在样品鉴定、采样及后期实验数据处理等方面提供的帮助, 感谢各位评审专家对文章的指导与修改。 |
Copy editor: 林强
收稿日期: 2022-06-17
修回日期: 2022-07-19
网络出版日期: 2022-07-26
基金资助
国家自然科学基金(41890853)
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0404)
国家科技基础资源调查专项(2018FY10010005)
中国科学院南海生态环境工程创新研究院自主部署项目(ISEE2018ZD02)
国家重点基础研究发展计划(2015CB452904)
国家重点研发计划(2017YFC0506302)
国家重点研发计划(2016YFC0502805)
Distribution characteristics and influencing factors of meso- and micro-zooplankton communities in the offshore waters of the Guangdong-Hong Kong-Macao Greater Bay Area*
Copy editor: LIN Qiang
Received date: 2022-06-17
Revised date: 2022-07-19
Online published: 2022-07-26
Supported by
National Nature Science Foundation(41890853)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0404)
National Science and Technology Fundamental Resources Investigation Program of China(2018FY10010005)
Key Project of Chinese National Programs for Fundamental Research and Development(2015CB452904)
National Key Research and Development Program of China(2017YFC0506302)
National Key Research and Development Program of China(2016YFC0502805)
Self-deployed project of Southern Marin Institute of Eco-Environmental Engineering Innovation, Chinese Academy of Sciences(NO.ISEE2018ZD02)
为阐明粤港澳大湾区近海水文动力和人为活动引起的环境变化对浮游动物群落结构的影响, 于2019年7月(夏季)和2020年12月(冬季)在粤港澳大湾区近海珠江口、大亚湾、黄茅海三个断面进行浮游动物采样, 比较分析中小型浮游动物的群落特征, 并探讨引起其季节和区域差异的影响因素。调查海域共鉴定中小型浮游动物211种(包括浮游幼虫), 其中桡足类占总种数的42.18%, 浮游动物的多样性、丰度呈现明显的季节和区域差异。浮游动物种数和平均丰度均为夏季高于冬季, 远岸海域物种多样性高于近岸。浮游动物丰度高值区在两个季节均位于珠江口咸淡水交汇区域; 夏季河口水域的平均丰度高于大亚湾水域的, 冬季丰度分布无明显差异。聚类分析结果显示, 调查海域夏季整体分为河口、海湾、远岸3个类群, 珠江口水域受径流影响较大, 与大亚湾之间群落结构差异明显; 而冬季河口与海湾近岸水域群落结构差异不明显。强额孔雀哲水蚤(Parvocalanus crassirostris)、短角长腹剑水蚤(Oithona brevicornis)在夏季和冬季均为优势种, 厦门矮隆哲水蚤(Bestiolina amoyensis)、中华异水蚤(Acartiella sinensis)、简长腹剑水蚤(Oithona simplex)等为夏季优势种, 小拟哲水蚤(Paracalanus parvus)等为冬季优势种。Pearson相关性分析与冗余分析表明大湾区近海浮游动物丰度与温度、盐度、叶绿素a浓度及营养物质的关系存在种间、季节和区域差异, 表明湾区复杂的水体环境条件对中小型浮游动物群落结构存在潜在影响。
宋星宇 , 林雅君 , 张良奎 , 向晨晖 , 黄亚东 , 郑传阳 . 粤港澳大湾区近海中小型浮游动物分布特征及影响因素*[J]. 热带海洋学报, 2023 , 42(3) : 136 -148 . DOI: 10.11978/2022137
In order to clarify the influence of hydrological dynamics and environmental changes caused by human activities on the zooplankton community structure in the coastal waters of Guangdong-Hong Kong-Macao Bay Area, zooplankton samples were collected in 3 transections (in the Pearl River Estuary, Daya Bay and Huangmao Sea) in July 2019 (summer) and December 2020 (winter). The characteristics of meso- and micro-zooplankton community were analyzed, and the influencing factors of their seasonal and regional differences were discussed. A total of 211 zooplankton species (including larvae) were identified in the study area, among which copepods accounted for 42.18%. The diversity and abundance of zooplankton showed obvious seasonal and regional differences. The number of species and average abundance of zooplankton were higher in summer than that in winter, and the species diversity in offshore waters was higher than that in nearshore waters. The high values of zooplankton abundance were located in the interface of saline- and plume- waters of the Pearl River Estuary in both seasons. In summer, the average abundance in estuarine waters was obviously higher than that in the Daya Bay, while in winter there was no such difference in the nearshore waters. The results of cluster analysis showed that the zooplankton community could be divided into three groups, i.e. the estuarine group, the bay group and the far-shore group in summer. The estuarine waters was greatly affected by runoff, and the community structure was significantly different from that of the Daya Bay. However, there was no significant difference in community structure between the estuarine and the bay in winter. Parvocalanus crassirostris and Oithona brevicornis were dominant species in both seasons, Bestiolina amoyensis, Acartiella sinensis and Oithona simplex were also dominant in the wet season, while Paracalanus parvus was also dominant in the dry season. Pearson correlation analysis and RDA analysis results showed that there were interspecific and seasonal differences in the correlations between the abundance of different species and temperature, salinity, phytoplankton biomass and nutrients, suggesting that the complex water environment in the Greater Bay Area had a potential influence on the differentiation of meso- and micro-zooplankton community structure.
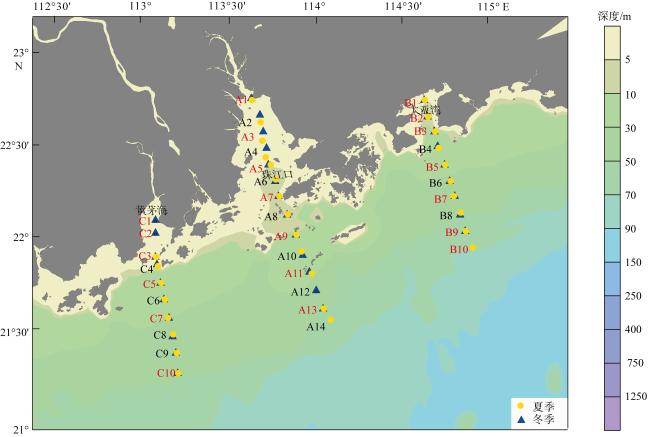
图1 粤港澳大湾区近海夏季和冬季浮游动物采样站位该图基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2019)4342的标准地图制作, 底图无修改。黄色圆点表示夏季调查站位, 蓝色三角形表示冬季调查站位, 红色站位标识表示浮游动物采样站位点 Fig.1 Zooplankton sampling sites in summer and winter in the Greater Bay Area offshore. Yellow dots indicate the summer survey sites, blue triangles indicate the winter survey sites, and red site signs mark the zooplankton sampling sites |
表1 大湾区近海不同类群浮游动物的种类数量Tab. 1 Number of zooplankton species in different groups in the Greater Bay Area offshore |
| 类群 | 夏季 | 冬季 | 共有种 | 合计 | 占比/% |
|---|---|---|---|---|---|
| 原生动物 | 1 | 1 | 0 | 2 | 0.95 |
| 轮虫 | 4 | 0 | 0 | 4 | 1.9 |
| 刺胞动物 | 32 | 15 | 11 | 36 | 17.06 |
| 栉水母 | 1 | 1 | 1 | 1 | 0.47 |
| 异足类 | 1 | 1 | 1 | 1 | 0.47 |
| 翼足类 | 4 | 3 | 3 | 4 | 1.9 |
| 枝角类 | 6 | 2 | 2 | 6 | 2.84 |
| 介形类 | 2 | 3 | 1 | 4 | 1.9 |
| 桡足类 | 80 | 62 | 53 | 89 | 42.18 |
| 糠虾类 | 1 | 3 | 0 | 4 | 1.9 |
| 端足类 | 4 | 4 | 1 | 7 | 3.32 |
| 磷虾类 | 1 | 1 | 0 | 2 | 0.95 |
| 十足类 | 2 | 3 | 2 | 3 | 1.42 |
| 涟虫 | 0 | 1 | 0 | 1 | 0.47 |
| 毛颚类 | 10 | 9 | 8 | 11 | 5.21 |
| 被囊类 | 9 | 7 | 5 | 11 | 5.21 |
| 浮游幼虫 | 21 | 20 | 16 | 25 | 11.85 |
| 合计 | 179 | 136 | 104 | 211 | 100 |
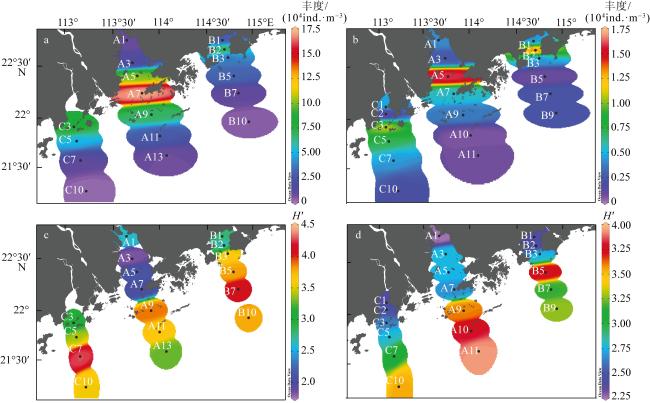
表2 大湾区近海浮游动物丰度和多样性特征Tab. 2 Abundance and diversity characteristics of zooplankton in the Greater Bay Area offshore |
| 季节 | 项目 | 平均值 | 范围 |
|---|---|---|---|
| 夏季 | 丰度/(104ind.·m-3) | 4.26±4.34 | 0.24~17.03 |
| 多样性指数(H′) | 3.21±0.73 | 1.83~4.28 | |
| 均匀度(J′) | 0.57±0.08 | 0.38~0.69 | |
| 冬季 | 丰度/(104ind.·m-3) | 0.48±0.43 | 0.05~1.59 |
| 多样性指数(H′) | 2.97±0.52 | 2.25~3.94 | |
| 均匀度(J′) | 0.58±0.07 | 0.49~0.72 |
表3 大湾区近海夏季和冬季浮游动物优势种Tab. 3 Dominant species of zooplankton in summer and winter off the Greater Bay Area |
| 浮游动物物种 | 平均丰度/(104ind.·m-3) | 占比/% | 优势度 | |
|---|---|---|---|---|
| 夏季 | 强额孔雀哲水蚤Parvocalanus crassirostris | 1.07±1.59 | 25.02 | 0.24 |
| 厦门矮隆哲水蚤Bestiolina amoyensis | 0.26±0.53 | 6.16 | 0.03 | |
| 中华异水蚤Acartiella sinensis | 0.24±0.57 | 5.59 | 0.02 | |
| 简长腹剑水蚤Oithona simplex | 0.21±0.36 | 4.86 | 0.03 | |
| 短角长腹剑水蚤Oithona brevicornis | 0.20±0.29 | 4.72 | 0.04 | |
| 冬季 | 强额孔雀哲水蚤Parvocalanus crassirostris | 0.09±0.15 | 19.28 | 0.18 |
| 短角长腹剑水蚤Oithona brevicornis | 0.03±0.07 | 6.96 | 0.07 | |
| 小拟哲水蚤Paracalanus parvus | 0.03±0.04 | 5.44 | 0.05 |
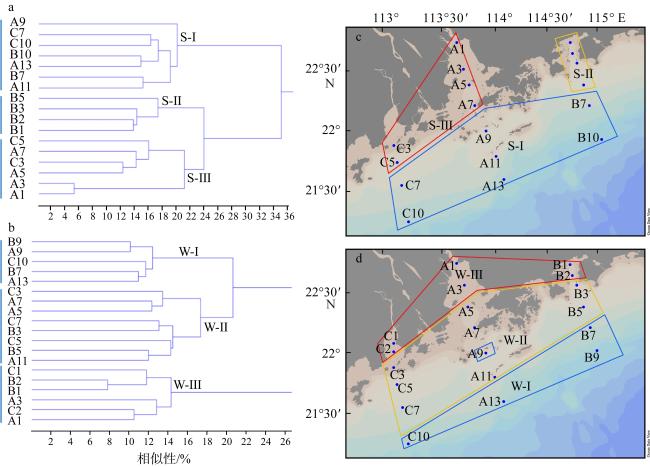
图3 大湾区近海夏季(a、c)和冬季(b、d)浮游动物群落分布聚类Fig. 3 Distribution and clustering of the zooplankton communities in summer (a, c) and winter (b, d) in the offshore of the Greater Bay Area |
表4 大湾区近海夏季和冬季不同浮游动物群落丰度、多样性及优势种Tab. 4 Abundance, diversity and dominant species of different zooplankton communities in the Greater Bay Area offshore in summer and winter |
| 夏季 | 冬季 | ||||||
|---|---|---|---|---|---|---|---|
| 群落S-I | 群落S-Ⅱ | 群落S-Ⅲ | 群落W-I | 群落W-Ⅱ | 群落W-Ⅲ | ||
| 丰度/(104 ind.·m-3) | 1.87±2.18a | 4.16±1.86ab | 7.13±5.53b | 0.23±0.09 | 0.61±0.49 | 0.51±0.44 | |
| 多样性指数(H') | 3.8±0.31a | 3.21±0.52a | 2.5±0.55b | 3.51±0.25a | 3.03±0.44b | 2.45±0.16c | |
| 均匀度指数(J') | 0.61±0.05a | 0.57±0.06ab | 0.51±0.08b | 0.62±0.05 | 0.58±0.08 | 0.54±0.05 | |
| 优势种 | 短角长腹剑水蚤(0.11) 强额孔雀哲水蚤(0.09) 肥胖三角溞(0.02) 挪威小毛猛水蚤(0.03) | 强额孔雀哲水蚤(0.29) 简长腹剑水蚤(0.14) 短角长腹剑水蚤(0.05) 肥胖三角溞(0.04) 鸟喙尖头溞(0.03) 尖额谐猛水蚤(0.03) | 强额孔雀哲水蚤(0.23) 厦门矮隆哲水蚤(0.07) 中华异水蚤(0.09) | 长尾住囊虫(0.08) 小拟哲水蚤(0.04) 平大眼剑水蚤(0.04) 尖额谐猛水蚤(0.03) 针刺拟哲水蚤(0.03) 挪威小毛猛水蚤(0.02) | 强额孔雀哲水蚤(0.14) 小拟哲水蚤(0.07) 短角长腹剑水蚤(0.03) | 强额孔雀哲水蚤(0.34) 短角长腹剑水蚤(0.15) 简长腹剑水蚤(0.04) | |
注: 不同的字母表示群落之间存在显著性差异(P< 0.05); 各物种后的数值为该物种的优势度 |
表5 大湾区近海夏季和冬季不同浮游动物群落所处区域的水体环境特征Tab. 5 Characteristics of the water environment in the areas where different zooplankton communities are located in the Greater Bay Area offshore in summer and winter |
| 环境特征 | 夏季表层 | 夏季底层 | 冬季表层 | 冬季底层 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 群落S-I | 群落S-Ⅱ | 群落S-Ⅲ | 群落S-I | 群落S-Ⅱ | 群落S-Ⅲ | 群落W-I | 群落W-Ⅱ | 群落W-Ⅲ | 群落W-I | 群落W-Ⅱ | 群落W-Ⅲ | |||
| 温度/℃ | 30.21±0.67a | 28.44±0.7b | 28.93±1.47ab | 22.86±0.61a | 23.51±1.08a | 25.76±1.42b | 20.92±0.72a | 20.05±0.58a | 19.05±1.01b | 21.49±0.88a | 20.24±0.69b | 19.25±0.75c | ||
| 盐度/‰ | 28.42±4.79a | 32.71±0.44a | 3.59±3.4b | 34.64±0.46 | 34.33±0.2 | 25±12.02 | 32.09±0.53a | 31±1.22a | 21.64±8.23b | 32.62±0.65a | 31.21±1.03a | 24.16±6.72b | ||
| pH | 8.51±0.16 | 8.41±0.03 | 8.11±0.52 | 8.27±0.06a | 8.1±0.11ab | 8.01±0.18b | 7.92±0.08 | 7.8±0.17 | 7.75±0.14 | 8.03±0.02a | 7.95±0.05a | 7.82±0.13b | ||
| DO/(mg·L-1) | 7.93±1.51 | 7.02±0.17 | 8.08±2.59 | 5.56±0.58 | 3.69±0.33 | 5.1±2 | 7.57±0.25a | 7.66±0.29a | 8.33±0.82b | 7.1±0.18 | 7.17±0.11 | 7.44±0.5 | ||
| 浊度/FNU | 0.57±0.58a | 2.49±2.01ab | 11.46±13.15b | 1.41±1.44a | 3.62±1.05a | 17.5±11.69b | 1.43±1.61a | 7.89±8.46ab | 21.4±19.51b | 1.06±0.54a | 18.97±25.53ab | 37.05±34.83b | ||
| DIN/(μmol·L-1) | 16.36±24.08a | 2.17±0.18a | 166.67±54.97b | 5.82±2.78a | 6.08±2.49a | 72.58±55.56b | 12.16±2.74 | 20.77±10.99 | 50.77±49.77 | 10.69±5.55a | 17.21±7.36a | 49.33±43.91b | ||
| DIP/(μmol·L-1) | 0.44±0.1a | 0.56±0.16a | 2.67±1.34b | 1.05±0.23a | 1.1±0.14a | 2.69±0.92b | 0.4±0.08a | 0.48±0.25a | 0.87±0.37b | 0.46±0.23 | 0.58±0.22 | 0.78±0.28 | ||
| N:P | 31.83±44.53ab | 4.12±0.99a | 82.97±49.08b | 5.36±1.99a | 5.38±1.94a | 24.25±16.18b | 32.49±10.83 | 134.57±213.8 | 51.67±37.21 | 34.39±29.06 | 29.93±43.88 | 52.44±38.91 | ||
| SiO2- 3/(μmol·L-1) | 25.28±27.55a | 15.84±4.34a | 162.06±38.14b | 16.89±6.48a | 22.12±11.82a | 106.29±63.93b | 33.46±2.43a | 49.99±8.74a | 102.86±53.2b | 31.24±7.97a | 46.18±10.64a | 106.61±50.77b | ||
| Chl a/(μg·L-1) | 5.25±7.17 | 3.29±2.07 | 5.59±5.22 | 1.41±1.25 | 3.13±1.89 | 2.08±0.99 | 1.28±0.29a | 1.59±0.57a | 2.67±0.86b | 1.27±0.54a | 1.54±0.44a | 2.28±0.63b | ||
注: 不同的字母表示群落之间存在显著差异; DO: 溶解氧; DIN: 溶解性无机氮; DIP: 溶解性无机磷; SiO2- 3: 硅酸盐 |
表6 大湾区近海夏季和冬季浮游动物丰度和多样性指数与表、底层环境因子的Pearson相关性Tab. 6 Pearson correlation of zooplankton abundance and diversity indices with surface and bottom layers environmental factors in the Greater Bay Area offshore in summer and winter |
| 丰度 | H' | 温度 | 盐度 | pH | DO | 浊度 | DIP | SiO2- 3 | DIN | N:P | Chl a | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 夏季 | 丰度 | 表层 | - | -0.51* | 0.021 | -0.472 | -0.095 | 0.118 | 0.1 | 0.511* | 0.53* | 0.404 | 0.328 | 0.254 |
| 底层 | -0.51* | 0.41 | 0.027 | -0.239 | -0.412 | 0.283 | 0.284 | 0.05 | -0.002 | 0.024 | 0.23 | |||
| H' | 表层 | -0.51* -0.51* | - | 0.492* | 0.701** | 0.583* | 0.298 | -0.635** | -0.813** | -0.77** | -0.718** | -0.195 | 0.116 | |
| 底层 | -0.778** | 0.685** | 0.792** | 0.203 | -0.8** | -0.752** | -0.707** | -0.684** | -0.649** | -0.302 | ||||
| 冬季 | 丰度 | 表层 | - | -0.484* -0.484* | -0.162 | 0.073 | 0.184 | -0.123 | 0.102 | -0.029 | -0.019 | -0.001 | 0.054 | 0.33 |
| 底层 | -0.299 | 0.021 | -0.057 | 0.052 | 0.061 | 0.249 | 0.003 | -0.054 | -0.304 | 0.509* | ||||
| H' | 表层 | -0.484* -0.484* | - | 0.667** | 0.551* | 0.178 | -0.403 | -0.388 | -0.401 | -0.572* | -0.422 | -0.175 | -0.637** | |
| 底层 | 0.733** | 0.56* | 0.608 | -0.384 | -0.45 | -0.686** | -0.613** | -0.479* | 0.123 | -0.735** | ||||
注: DO: 溶解氧; DIP: 溶解性无机磷; SiO2- 3: 硅酸盐; DIN: 溶解性无机氮; *表示显著相关(P<0.05); **表示极显著相关(P<0.01) |
| [1] |
陈菊芳, 齐雨藻, 徐宁, 等, 2005. 大亚湾拟菱形藻水华及其在生物群落中的生态地位[J]. 海洋学报(中文版), 27(1): 114-119.
|
| [2] |
陈清潮, 章淑珍, 1974. 南海浮游桡足类I[J]. 海洋科学集刊, 9: 101-124.
|
| [3] |
丁峰元, 李圣法, 董婧, 等, 2005. 春季东海区近海浮游动物群落结构及其影响因子[J]. 海洋渔业, 27(1): 26-32.
|
| [4] |
高原, 赖子尼, 庞世勋, 等, 2010. 黄茅海河口虎跳门和崖门浮游动物群落的比较[J]. 生态科学, 29(2): 121-126.
|
| [5] |
郭沛涌, 沈焕庭, 刘阿成, 等, 2008. 长江河口中小型浮游动物数量分布、变动及主要影响因素[J]. 生态学报, 28(8): 3517-3525.
|
| [6] |
黄彬彬, 郑淑娴, 蔡伟叙, 等, 2017. 珠江口枯水期和丰水期中小型桡足类种类组成、丰度分布及其与环境因子的关系[J]. 厦门大学学报(自然科学版), 56(6): 852-858.
|
| [7] |
黄彬彬, 郑淑娴, 田丰歌, 2019. 珠江口枯水期和丰水期中小型浮游动物群落动态[J]. 应用海洋学学报, 38(1): 43-52.
|
| [8] |
黄道建, 郭振仁, 陈菊芳, 等, 2010. 大亚湾代表水域浮游动物生物量的垂直分布与季节动态研究[J]. 海洋环境科学, 29(6): 825-828.
|
| [9] |
黄道建, 于锡军, 郭振仁, 等, 2009. 珠江口黄茅海冬季海洋生态环境特征[J]. 海洋通报, 28(2): 28-33.
|
| [10] |
黄加祺, 1983. 九龙江口大、中型浮游动物的种类组成和分布[J]. 厦门大学学报(自然科学版), 22(1): 88-95.
|
| [11] |
黄小平, 2019. 营养物质对海湾生态环境影响的过程与机理[M]. 北京: 科学出版社, 1-399.
|
| [12] |
黄亚如, 欧强, 李锦蓉, 等, 1993. 南海大鹏湾与大亚湾浮游动物的比较及其与赤潮的关系[J]. 海洋通报, 12(2): 46-51.
|
| [13] |
李开枝, 尹健强, 黄良民, 等, 2005. 珠江口浮游动物的群落动态及数量变化[J]. 热带海洋学报, 24(5): 60-68.
|
| [14] |
李开枝, 尹健强, 黄良民, 等, 2007. 珠江口浮游桡足类的生态研究[J]. 生态科学, (2): 97-102.
|
| [15] |
李晓慧, 刘镇盛, 2017. 长江口及邻近海域浮游动物生物量分布及季节变化[J]. 海洋学研究, 35(4): 94-101.
|
| [16] |
李优迈, 韩留玉, 陈绵润, 等, 2018. 大亚湾中型浮游动物群落结构和植食性[J]. 海洋与湖沼, 49(4): 839-850.
|
| [17] |
连喜平, 谭烨辉, 黄良民, 等, 2011. 大亚湾大中型浮游动物的时空变化及其影响因素[J]. 海洋环境科学, 30(5): 640-645.
|
| [18] |
刘华健, 黄良民, 谭烨辉, 等, 2017. 珠江口浮游植物叶绿素a和初级生产力的季节变化及其影响因素[J]. 热带海洋学报, 36(1): 81-91.
|
| [19] |
刘镇盛, 2012. 长江口及其邻近海域浮游动物群落结构和多样性研究[D]. 青岛: 中国海洋大学, 116-118.
|
| [20] |
马婕, 2019. 大亚湾浮游动物主要功能群的摄食及其对环境变化的响应[D]. 上海: 上海海洋大学, 31-41.
|
| [21] |
孙翠慈, 王友绍, 孙松, 等, 2006. 大亚湾浮游植物群落特征[J]. 生态学报, 26(12): 3948-3958.
|
| [22] |
文小明, 刘佳, 陈传忠, 等, 2021. 粤港澳大湾区生态环境监测发展现状与展望[J]. 中国环境监测, 37(5): 14-20.
|
| [23] |
谢福武, 梁计林, 邢孔敏, 2019. 海南岛近岸海域夏、秋季浮游动物分布特征研究[J]. 海岸工程, 38(4): 280-293.
|
| [24] |
徐兆礼, 陈亚瞿, 1989. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J]. 生态学杂志, 8(4): 13-15.
|
| [25] |
杨威, 董园, 俎婷婷, 等, 2019. 南海北部夏季叶绿素a分布规律及影响因素[J]. 热带海洋学报, 38(6): 9-20.
|
| [26] |
尹天齐, 王庆, 杨宇峰, 等, 2022. 基于形态学和DNA分子鉴定的珠江口浮游动物群落结构比较研究[J]. 热带海洋学报, 41(3): 172-185.
|
| [27] |
袁丹妮, 2014. 珠江口广州—珠海水域浮游动物群落结构及其环境特征[D]. 广州: 暨南大学, 34-56.
|
| [28] |
张达娟, 闫启仑, 王真良, 2008. 典型河口浮游动物种类数及生物量变化趋势的研究[J]. 海洋与湖沼, 39(5): 536-540.
|
| [29] |
张立明, 谭烨辉, 李佳俊, 等, 2020. 大亚湾夏季浮游植物群落结构及对淡澳河输入的响应特征[J]. 热带海洋学报, 39(5): 43-54.
|
| [30] |
张良奎, 林雅君, 向晨晖, 等, 2022. 粤港澳大湾区近海浮游植物群落结构特征及其驱动因素[J]. 生态科学(已录用).
|
| [31] |
张武昌, 高尚武, 孙军, 等, 2010. 南海北部冬季和夏季浮游哲水蚤类群落[J]. 海洋与湖沼, 41(3): 448-458.
|
| [32] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008a. GB/T 12763. 4-2007 海洋调查规范第4部分:海水化学要素调查[S]. 北京, 中国标准出版社.
GENERAL ADMINISTRATION OF QUALITY SUPERVISION, INSPECTION AND QUARANTINE OF THE PEOPLE'S REPUBLIC OF CHINA, STANDARDIZATION ADMINISTRATION, 2008. GB/T 12763. 4-2007 Specifications for oceanographic survey-Part 4: Survey of chemical parameters in sea water[S]. Beijing: Standards Press of China. (in Chinese)
|
| [33] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008b. GB/T 12763. 6-2007 海洋调查规范第6部分:海洋生物调查[S]. 北京, 中国标准出版社.
GENERAL ADMINISTRATION OF QUALITY SUPERVISION, INSPECTION AND QUARANTINE OF THE PEOPLE'S REPUBLIC OF CHINA, STANDARDIZATION ADMINISTRATION, 2008. GB/T 12763. 6-2007 Specifications for oceanographic survey-Part 6: Survey of marine organism[S]. Beijing: Standards Press of China. (in Chinese)
|
| [34] |
邹清, 姚炜民, 张淑敏, 2012. 乐清湾2009年夏季浮游动物种类组成和数量分布[J]. 海洋科学进展, 30(2): 266-275.
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
SCHLITZER, REINER, 2020. Ocean Data View[DB/OL]. [2022-06-17]. https://odv.awi.de.
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}