
2019年春季孟加拉湾中部放射虫群落结构垂向变化*
|
张兰兰(1978—), 女, 山东省青州人, 研究员, 主要从事海洋微体古生物现代过程与古海洋学研究。email: |
Copy editor: 姚衍桃
收稿日期: 2022-09-30
修回日期: 2022-11-09
网络出版日期: 2022-11-10
基金资助
国家自然科学基金面上项目(41876056)
国家自然科学基金面上项目(42176080)
中国科学院南海海洋研究所自主部署项目(SCSIO202201)
国家自然科学基金委员会共享航次计划项目(41849910)
Changes of radiolarian community structure with depth in the central Bay of Bengal in spring 2019
Copy editor: YAO Yantao
Received date: 2022-09-30
Revised date: 2022-11-09
Online published: 2022-11-10
Supported by
National Natural Science Foundation of China(41876056)
National Natural Science Foundation of China(42176080)
Development fund of South China Sea Institute of Oceanology of the Chinese Academy of Sciences(SCSIO202201)
National Natural Science Foundation of China Shiptime Sharing Project(41849910)
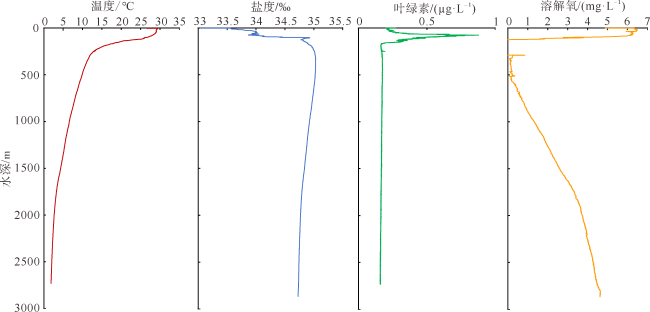
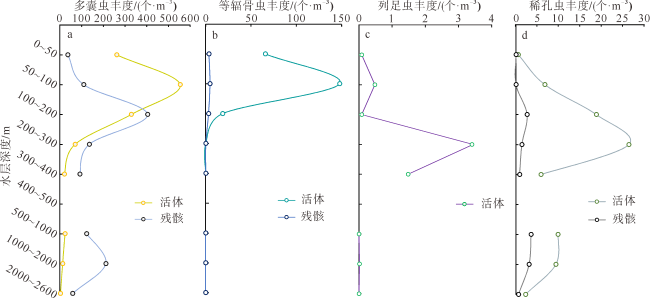
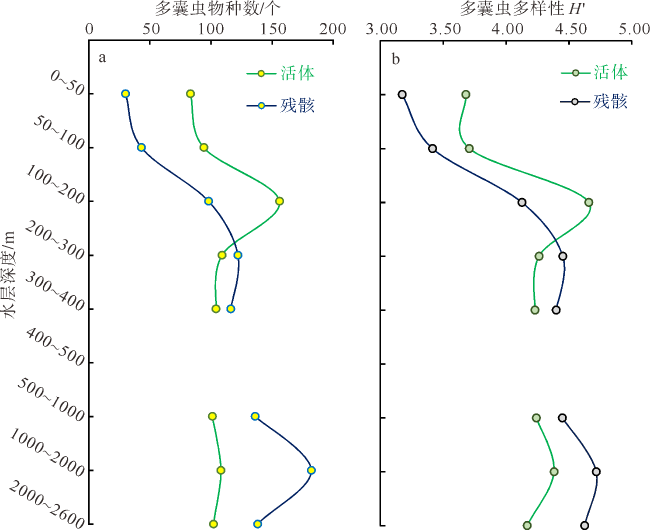
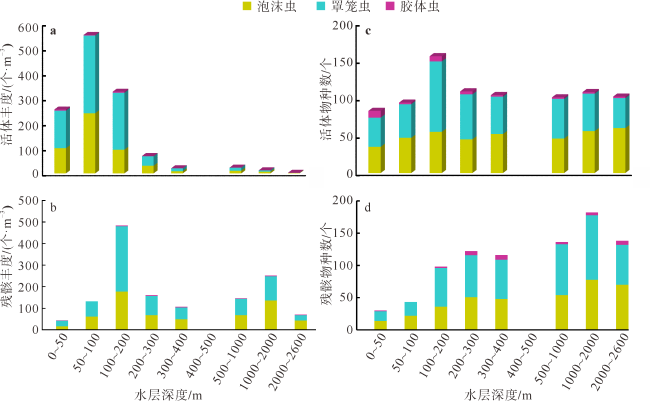
本文利用虎红染色和传统形态分析方法, 首次揭示了孟加拉湾中部0~2600m水深放射虫类群(多囊虫、等辐骨虫、列足虫和稀孔虫)的垂向变化特征。多囊虫可生活于全水深, 丰度最高值在叶绿素最大值层, 表明大部分多囊虫偏好光线充足和营养丰富的水体环境; 在阿拉伯海高盐水入侵影响的中深层水体(200~1000m)中, 多囊虫活体群和残骸群的丰度变化平稳, 反映出其偏好高盐环境(>34.5‰); 1000~2000m水层中活体群未增加, 但其残骸群的丰度明显升高, 推测取样期间1000~2000m水深处可能存在横向搬运现象。等辐骨虫活体丰度最高值出现在50~100m, 100~200m深度急剧下降, 在200~2600m水深则几乎缺失; 0~200m残骸体数量极低, 表明等辐骨虫偏好光线充足、营养丰富的环境, 其在约100m深发生大量溶解。印度棒矛虫(新种)(Sticholonche indicum sp. nov.)被描记, 其形态不同于其他棒矛虫种, 具有明显更长的船桨状轴足, 其活体峰值位于200~300m, 偏好中等温度、高盐的环境, 且不受低含氧量的限制。稀孔虫主要生活于50m以深, 高峰值出现在200~300m, 300~2600m少量存在, 其残骸群在0~2600m都处于极低值, 反映出低氧含量对稀孔虫影响不明显, 而且部分稀孔虫物种不受低温限制, 可生存于深水中。可见, 浮游动物多囊虫、等辐骨虫、列足虫和稀孔虫具有显著不同的垂直分布与环境适应特征, 本研究有助于理解海洋生态系统的物质循环和环境变化。
张兰兰 , 程夏雯 , 向荣 , 邱卓雅 , 常虎 . 2019年春季孟加拉湾中部放射虫群落结构垂向变化*[J]. 热带海洋学报, 2023 , 42(4) : 166 -175 . DOI: 10.11978/2022209
By staining with Rose Bengal and using traditional morphological analysis, this study for the first time reveals the vertical changes of radiolarian community (Polycystinea, Acanthadaria, Taxopodia and Phaeodaria) from 0 to 2600m water depth in the central Bay of Bengal. Polycystinea may survive in full water depth, and the highest abundance is in the chlorophyll-a maximum layer, indicating that most Polycystinea prefers the well-lit and nutrient-rich environments. In the intermediate-deep water (200~1000 m) invaded by high saline water from the Arabian Sea, the change of abundances of living and dead Polycystinea are slight, indicating that they prefer high-salinity environment (>34.5‰). Considering that living Polycystinea did not change significantly from 1000m to 2000m, the abundance of the shells was obviously increased compared with that in the upper water, indicating that the central Bay of Bengal was affected by the lateral advection of ocean current in the 1000~2000m during the sampling period. The highest abundance of Acantharia appeared at 50~100m, then decreased sharply at 100~200m depth, and almost absent at 200~2600m. The number of shells from 0 to 200m was low, suggesting that Acantharia prefers the well-lit and nutrient-rich environments, and is speculated to be largely dissolved at ~100m depth. Different from other species in morphology, a new species Sticholonche indicum sp. nov. was described, which has the significantly longer oar-like axopodia. And its maximum abundance occurs between 200~300m, suggesting that they are new species, which prefer the moderate temperature and high salt environment, not limited by the oxygen minimum zone (OMZ). Phaeodaria mainly lives at a depth of 50m, with the highest abundance at 200~300m depth, the low abundance at 300~2600m depth, and the rare shells of thanatocoenoses at 0~2600m depth, suggesting that Phaeodaria is weakly affected by the OMZ, and some phaeodarian species can live in deep water without the limit of low temperature. Therefore, Polycystinea, Acantharia, Taxopodia and Phaeodaria have the significant differences in vertical distribution and environmental adaptation characteristics, and the above results would be helpful to understand the material cycle and environmental changes in marine ecosystems.
Key words: Bay of Bengal; zooplankton radiolaria; community structure; depth change
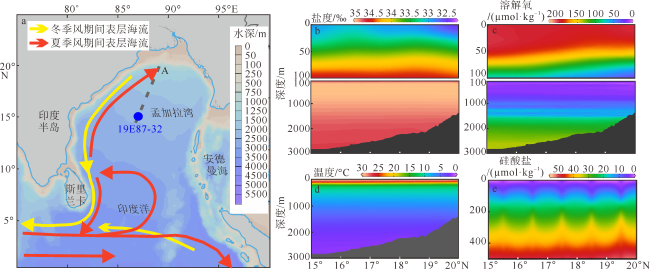
图1 研究站位(19E87-32)的地理位置与断面A环境参数剖面图图a Locations of multi-network sampling station (19E87-32) in the Bay of Bengal, and Section A profile of environmental parameters. (a) is based on the standard map No. GS(2016)1663 downloaded from the standard map service website of the State Bureau of Surveying and Mapping; (b), (c), (d) and (e) are the quarterly mean salinity, dissolved oxygen, temperature and silicate during April-June from 1955 to 2017 with depth for Section A. Data are from https://odv.awi.de/data/ocean/world-ocean-atlas-2018 |
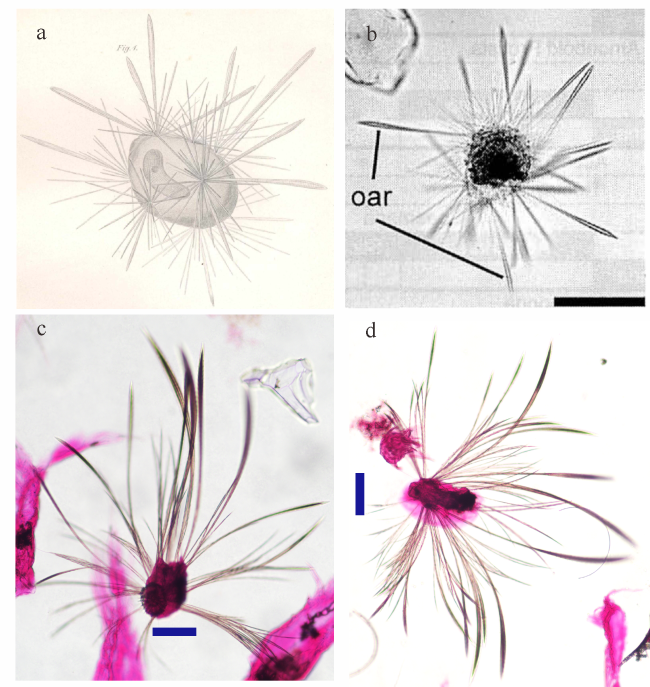
图4 列足虫物种形态对比a. 镰形棒矛虫(Sticholonche zanclea)(Fol, 1883)(模式标本); b. 镰形棒矛虫(Sticholonche zanclea)(鈴木紀毅 等, 2011); c. 印度棒矛虫(Sticholonche indicum sp. nov.)(模式标本); d. 印度棒矛虫。图中标尺长度为100μm Fig.4 Comparison of Taxopodia species morphology. (a) Sticholonche zanclea (Fol, 1883) (type specimen); (b) Sticholonche zanclea (Suzuki et al, 2011); (c, d) Sticholonche indicum sp. nov. (c for type specimen) |
*感谢中国科学院中国-斯里兰卡联合科教中心的资助; 感谢自然资源部海洋生态系统动力学重点实验室开放研究基金资助; 感谢“实验3”号科考船全体队员的帮助; 感谢日本东北大学Noritoshi Suzuki教授和3位评审专家的建设性意见和建议。
| [1] |
胡维芬, 张兰兰, 陈木宏, 等, 2015. 南海断面春季活体放射虫生态分布及其对环境的响应[J]. 中国科学: 地球科学, 45(1): 83-98.
|
| [2] |
鈴木紀毅, 相田吉昭, 2011. 放散虫の生物学一分布, 现存量, 共生生物[J]. 日本プランクト学会, 58(1): 40-48.
|
| [3] |
谭智源, 高洪绪, 宿星慧, 1978. 镰形棒矛虫在东海西部的数量分布[J]. 海洋与湖沼, 9(1): 59-66.
|
| [4] |
张杰, 张兰兰, 陈木宏, 等, 2020. 现代放射虫的高阶分类现状及其生态学意义[J]. 微体古生物学报, 37(1): 82-98.
|
| [5] |
张兰兰, 邱卓雅, 向荣, 等, 2021. 基于生物硅记录的孟加拉湾东南部末次冰期以来的古生产力变化[J]. 地学前缘, 29(4): 136-143.
|
| [6] |
张武昌, 张翠霞, 肖天, 2009. 海洋浮游生态系统中小型浮游动物的生态功能[J]. 地球科学进展, 24(11): 1195-1201.
|
| [7] |
张作人, 谭智源, 1964. 东海放射虫的研究 Ⅰ. 等辐骨虫目[J]. 海洋科学集刊, 6: 33-78.
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}