
基于线粒体控制区的中国近海竹荚鱼种群遗传结构和种群历史动态分析
|
任慧敏(1995—), 女, 贵州省铜仁市人, 硕士研究生, 从事海洋分子生态学研究。email: |
Copy editor: 殷波
收稿日期: 2020-11-02
要求修回日期: 2021-02-22
网络出版日期: 2021-02-25
基金资助
国家重点研发计划项目(2018YFD0900902)
国家重点研发计划项目(2018YFD0900904)
中国科学院战略性先导科技专项(B类)(XDB42030203)
上海科委“科技创新行动计划”“一带一路”国际合作项目(18230743200)
版权
Population genetic structure and historical dynamics of Trachurus japonicus in the China seas based on mitochondrial control region
Copy editor: YIN Bo
Received date: 2020-11-02
Request revised date: 2021-02-22
Online published: 2021-02-25
Supported by
National Key Research and Development Program of China(2018YFD0900902)
National Key Research and Development Program of China(2018YFD0900904)
Strategic Priority Research Program of Chinese Academy of Sciences(XDB42030203)
Shanghai Science and Technology Committee “Science and Technology Innovation Action Plan”“One Belt and One Road” International Cooperation Project of Shanghai(18230743200)
Copyright
竹荚鱼(Trachurus japonicus)是中国近海主捕鱼种之一, 在海洋食物网中扮演了重要角色, 然而环境污染和过度捕捞导致其出现种群数量衰退以及个体趋于小型化等现象。为了解中国近海竹荚鱼的种群遗传格局, 文章以线粒体DNA控制区为遗传标记研究了东海大陆架、福建近海和南海北部湾竹荚鱼群体的遗传结构以及种群历史动态。结果表明, 中国近海竹荚鱼整体呈现高单倍型多态性(Hd=0.998±0.001)和高核苷酸多态性(π=0.01259±0.00041)的遗传多样性特征。单倍型网络图呈现为星形辐射状的单一谱系, 利用最大似然法构建的系统进化树也未发现谱系分化。不同海区地理群体的分子方差分析显示东海群体和南海北部湾群体间无遗传分化, 遗传变异主要来源于群体内部(99.39%)。中性检验和核苷酸歧点分布分析结果暗示各海区竹荚鱼群体(东海、南海北部湾)以及整个群体均经历过近期的种群扩张。中国近海竹荚鱼群体呈现为遗传均匀的种群结构, 可以作为一个单一的种群加以管理, 人类高强度捕捞压力尚未影响其种群恢复潜力。
任慧敏 , 张衡 , 许莎莎 , 李圣法 , 李建生 , 李治洪 , 何利军 . 基于线粒体控制区的中国近海竹荚鱼种群遗传结构和种群历史动态分析[J]. 热带海洋学报, 2021 , 40(5) : 36 -44 . DOI: 10.11978/2020129
As a dominant fishing species, Trachurus japonicus plays an important role in marine food web. However, T. japonicus experienced rapid collapse in population size and decrease in body size due to environmental pollution and overfishing. To uncover evolutionary influence of anthropic activities on marine species in the China seas, we examined genetic structure and population historical dynamics of T. japonicus based on mitochondrial DNA control region. The results were shown in the following: 1) High haplotype diversity (Hd = 0.998±0.001) and high nucleotide diversity (π = 0.01259±0.00041) of T. japonicus were observed in the study areas; 2) A single star-like bursting lineage was identified in the reconstructed haplotypes network and phylogenetic tree based on maximum likelihood method; 3) Genetic variation of T. japonicus occurred mainly within the populations (99.39%) instead of among different geographic populations according to analyses of molecular variance, and no significant genetic differentiation was observed among the East China Sea shelf, coastal Fujian and Beibu Gulf populations; 4) Recent demographic expansions in the East China Sea shelf, coastal Fujian province, Beibu Gulf and overall populations were consistently suggested by neutral tests and mismatch distribution analysis. Thus, T. japonicus can be managed as a single and genetic homogeneous population in the China seas. Even if high fishing pressure had been kept on T. japonicus during the past several decades, strong recovery potential for this species could still be expected in the China seas.
表1 竹荚鱼取样信息及线粒体控制区序列遗传多态性参数Tab. 1 Sampling information and genetic diversity parameters of Trachurus japonicus based on mitochondrial control region |
| 海区 | 位置信息 | 序列来源 | Genbank登陆号 | 样本数 | 单倍型数 | 单倍型多样性 (Hd±SD) | 核苷酸多样性 (π±SD) |
|---|---|---|---|---|---|---|---|
| 东海 | 东海-大陆架 | 本研究 | / | 30 | 30 | 1.000±0.009 | 0.01170±0.00066 |
| 福建近海 | 牛素芳等(2011) | HM212543~HM212595 | 60 | 53 | 1.000±0.004 | 0.01288±0.00061 | |
| 合计 | / | / | 90 | 80 | 0.999±0.002 | 0.01248±0.00045 | |
| 南海-北部湾 | 南海-防城港 | Genbank | FJ914949~FJ914987 /GQ382142~GQ382149 | 47 | 43 | 0.996±0.005 | 0.01302±0.00078 |
| 中国近海 | 东海-南海 | / | / | 137 | 118 | 0.998±0.001 | 0.01259±0.00041 |
注: / 表示无此项 |
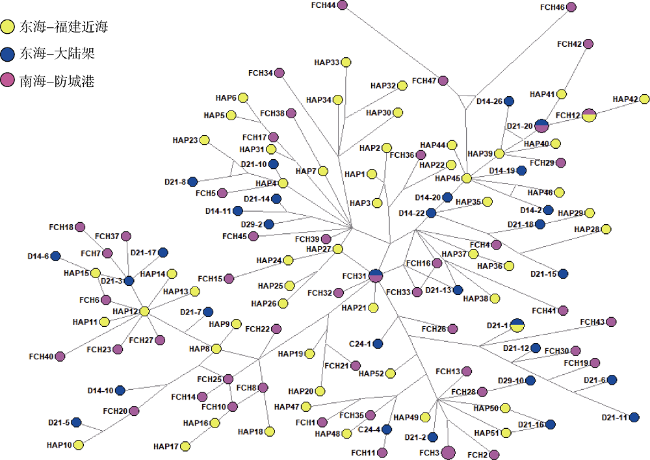
图1 基于线粒体控制区的竹荚鱼单倍型网络图不同颜色代表不同地理分布的单倍型。各单倍型地理信息为: C/D来自东海大陆架, HAP来自福建沿海, FCH来自南海北部湾防城港近岸 Fig. 1 Haplotype network of Trachurus japonicus based on mitochondrial control region. Geographic distribution information of each haplotype was distinguished using different colors: red ones from the Beibu Gulf in the South China Sea, blue ones from the East China Sea shelf, and yellow ones from coastal Fujian Province in the East China Sea |
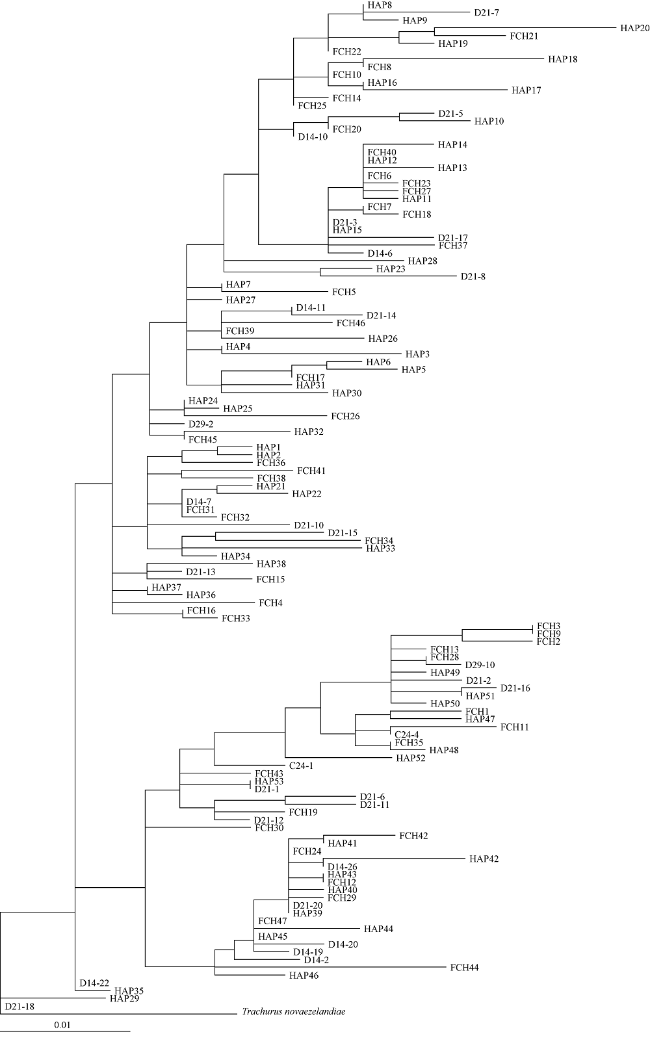
图2 基于线粒体控制区的竹荚鱼最大似然树各单倍型地理信息为: C/D来自东海大陆架, HAP来自福建沿海, FCH来自南海北部湾防城港近岸 Fig. 2 Maximum likelihood tree of Trachurus japonicus based on mitochondrial control region. Geographic information of each haplotype is indicated: C/D from the East China Sea shelf, HAP from coastal Fujian Provincce in the East China Sea, and FCH from coastal Fangchenggang City in the Beibu Gulf of the South China Sea |
表2 中国近海竹荚鱼群体的分子方差分析Tab. 2 AMOVA of Trachurus japonicus in the China seas based on mitochondrial control region |
| 变异来源 | 自由度 | 平方和 | 变异组成 | 变异百分比/% | 遗传分化系数(Fst) |
|---|---|---|---|---|---|
| 群体间 | 2 | 13.955 | 0.03385Va | 0.61 | / |
| 群体内 | 127 | 704.576 | 5.54784Vb | 99.39 | / |
| 整体 | 129 | 718.531 | 5.58169 | / | 0.00606 |
注: / 表示无此项; Va代表群体间变异; Vb代表群体内变异 |
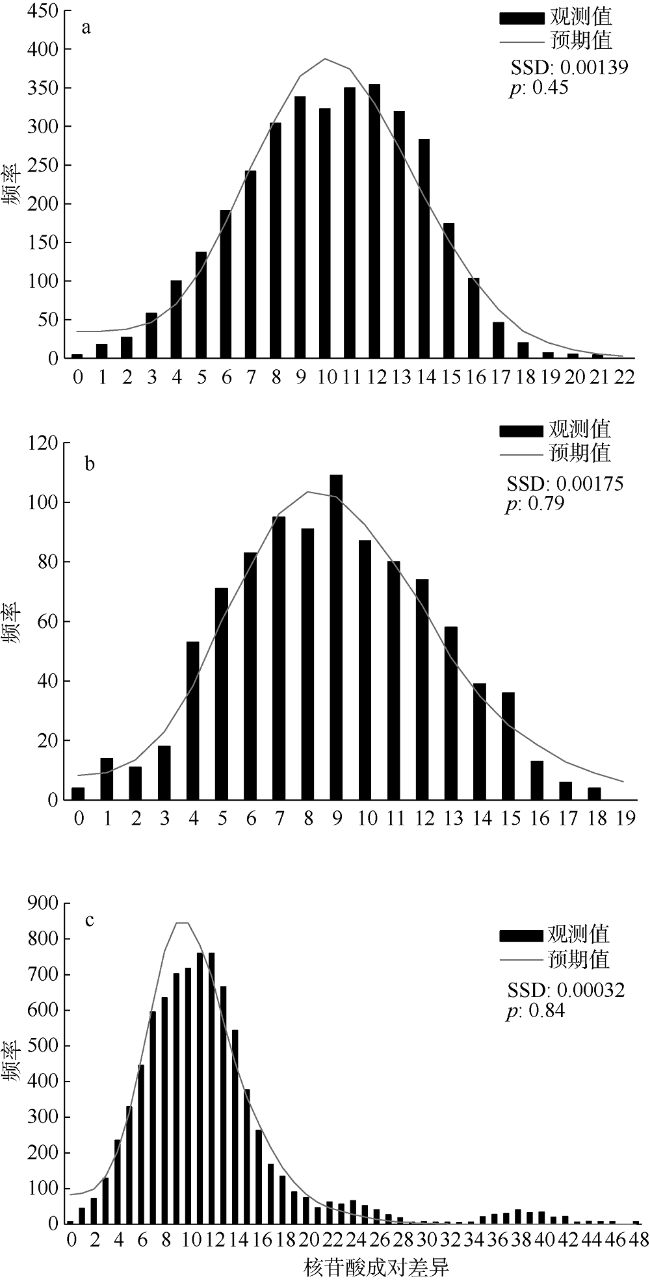
图3 竹荚鱼不同地理群体的单倍型歧点分布分析图a. 东海; b. 南海-北部湾; c. 整体。SSD表示离差平方和, p表示SSD统计的显著性值 Fig. 3 Mismatch distribution analysis of Trachurus japonicus |
表3 中国近海竹荚鱼群体的中性检验Tab. 3 Neutrality tests of Trachurus japonicus populations in the China seas |
| 群体 | 群体大小 | Fu’s Fs检验 | Tajima’s D检验 |
|---|---|---|---|
| 东海 | 83 | -24.506** | -1.111 |
| 南海-北部湾 | 47 | -24.302** | -1.319 |
| 整体 | 130 | -24.572** | -1.327 |
注: ** 表示差异极显著(p<0.01) |
| [1] |
滨田律子, 马永钧, 1989. 东海竹(竹夹)鱼渔获量的变动[J]. 国外水产, (1):40-42 (in Chinese).
|
| [2] |
曹宁, 高健, 2006. 东海竹荚鱼的开发利用和区域共同管理探讨[J]. 渔业经济研究, (5):25-29.
|
| [3] |
刘舜斌, 1992. 东海竹荚鱼资源数量变动及现状[J]. 海洋渔业, (1):41-43 (in Chinese).
|
| [4] |
马克平, 1993. 试论生物多样性的概念[J]. 生物多样性, 1(1):20-22.
|
| [5] |
牛素芳, 苏永全, 王军, 等, 2011. 福建近海竹荚鱼线粒体DNA控制区和细胞色素b遗传多态性[J]. 中国水产科学, 18(1):66-74.
|
| [6] |
萨姆布鲁克 J, 2009. 分子克隆实验指南 3版[M]. 北京: 科学出版社.
|
| [7] |
邵帼瑛, 张敏, 2006. 东南太平洋智利竹荚鱼渔场分布及其与海表温关系的研究[J]. 上海海洋大学学报, 15(4):468-472.
|
| [8] |
《世界大洋性渔业概况》编写组, 2011. 世界大洋性渔业概况[M]. 北京: 海洋出版社.
COMPILATION OF WORD OCEANIC FISHERY STATUS, 2011. World Oceanic Fishery Status[M]. Beijing: China Ocean Press (in Chinese).
|
| [9] |
唐涛, 周明成, 2005. 动物辞典下[M]. 呼和浩特: 远方出版社: 245-246(in Chinese).
|
| [10] |
张丽艳, 牛素芳, 张曼, 等, 2014. 基于AFLP分子标记探讨福建近海竹荚鱼群体遗传多样性[J]. 厦门大学学报(自然科学版), 53(1):120-125.
|
| [11] |
张秋华, 程家骅, 徐汉祥, 等, 2007. 东海区渔业资源及其可持续利用[M]. 上海: 复旦大学出版社: 256-263(in Chinese).
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}