
不同绿藻对模拟酸雨胁迫的生理生化响应特征
|
杜响(1997—), 男, 安徽省阜阳市人, 硕士研究生, 从事藻类生理生化研究。email: dxiang231023@163.com |
收稿日期: 2022-11-14
修回日期: 2023-01-07
网络出版日期: 2023-01-10
基金资助
现代农业产业技术体系专项资金项目(CARS-50)
南麂列岛国家海洋自然保护区管理局项目(H202200074)
Physiological and biochemical responses of different species of chlorophyta to simulated acid rain stress
Received date: 2022-11-14
Revised date: 2023-01-07
Online published: 2023-01-10
Supported by
China Agriculture Research System(CARS-50)
Nanji Islands National Marine Nature Reserve Administration Project(H202200074)
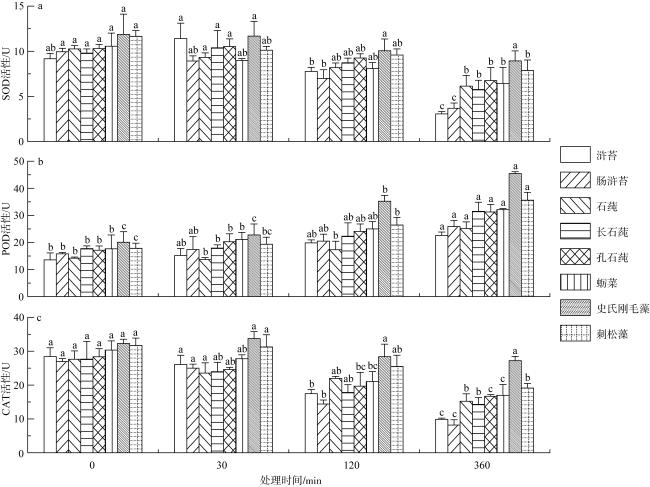
以东海潮间带绿藻优势种为试验材料, 测定其最大光化学量子产量(maximal photochemical quantum yield of PSⅡ, Fv/Fm)、可溶性糖含量及抗氧化酶活性等指标, 研究不同强度模拟酸雨(pH分别为6.3、5.6、4.5、4.0、3.5)胁迫下绿藻优势种生理生化响应。结果表明: 1) 随着模拟酸雨酸度和处理时间增加, 2种浒苔、4种石莼、史氏刚毛藻和刺松藻的Fv/Fm抑制幅度增大; 2) pH 4.5的模拟酸雨降低藻体中叶绿素a、类胡萝卜素和可溶性糖含量, 降幅为浒苔>石莼>刺松藻>史氏刚毛藻。而可溶性蛋白含量有所增加, 且随时间延长愈加显著, 史氏刚毛藻增幅高于其他绿藻。3) pH 4.5的模拟酸雨抑制绿藻中超氧化物歧化酶(superoxide dismutase, SOD)、过氧化氢酶(catalase, CAT)活性, 但提高其过氧化物酶(peroxidase, POD)活性, 且处理时间与POD活性正相关, 与SOD、CAT活性负相关。研究表明pH 4.5能作为模拟酸雨胁迫潮间带绿藻优势种的试验条件, 史氏刚毛藻对模拟酸雨耐受性最佳, 石莼比浒苔具有更好的抗酸雨胁迫能力。
杜响 , 骆其君 , 陈海敏 . 不同绿藻对模拟酸雨胁迫的生理生化响应特征[J]. 热带海洋学报, 2023 , 42(5) : 115 -123 . DOI: 10.11978/2022243
The maximal photochemical quantum yield of PSⅡ (Fv/Fm), soluble sugar contents and antioxidant enzyme activity were determined by using dominant species of chlorophyta in the intertidal zone of the East China Sea. The physiological and biochemical responses of dominant species of chlorophyta under simulated acid rain (pH 6.3, 5.6, 4.5, 4.0, 3.5, respectively) were studied. The results showed that: 1) the simulated acid rain inhibited Fv/Fm of Ulva prolifera and Ulva intestinalis, the remaining green seaweeds of Ulva, Cladophora stimpsonii and Codium fragile, and the inhibition amplitude increased with the increase of acidity and treatment time; 2) simulated acid rain at pH 4.5 reduced the contents of chlorophyll a, carotenoids and soluble sugar, and the decrease amplitude was green seaweeds of Ulva > C. fragile > C. stimpsonii. but the contents of soluble protein increased, which became more significant as time went on, and the increase amplitude of C. stimpsonii was higher than that of other green seaweeds; 3) simulated acid rain at pH 4.5 inhibited the activities of superoxide dismutase (SOD) and catalase (CAT), but increased the activities of peroxidase (POD) of seaweeds, and the treatment time was positively correlated with the activities of POD, but negatively correlated with the activities of SOD and CAT. In summary, pH 4.5 can be used as physiological and biochemical test conditions for dominant species of chlorophyta to simulated acid rain stress, C. stimpsonii exhibited higher tolerance to simulated acid rain, compared with U. prolifera and U. intestinalis, the remaining green seaweeds of Ulva has higher tolerance to simulated acid rain.
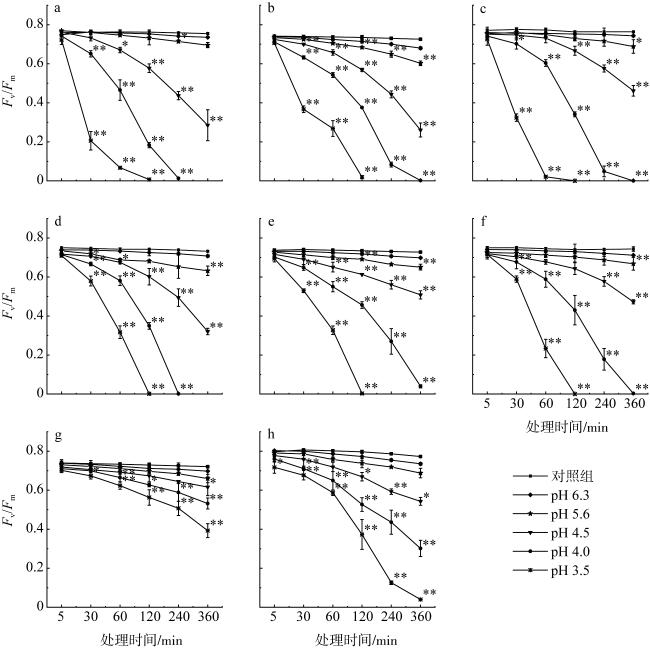
图1 不同pH值的模拟酸雨胁迫对8种绿藻Fv/Fm的影响a. 浒苔; b. 肠浒苔; c. 石莼; d. 长石莼; e. 孔石莼; f. 蛎菜; g. 史氏刚毛藻; h. 刺松藻。*和**表示在0.05和0.01水平上不同处理间差异显著 Fig. 1 Effects of simulated acid rain stress on Fv/Fm at different pH levels of eight species of green seaweeds. (a) U. prolifera; (b) U. indicate; (c) U. lactuca; (d) U. stenophylla; (e) U. pertusa; (f) U. conglobata; (g) C. stimpsonii; (h) C. fragile. * and ** indicate significant difference among different treatments at 0.05 and 0.01 levels |
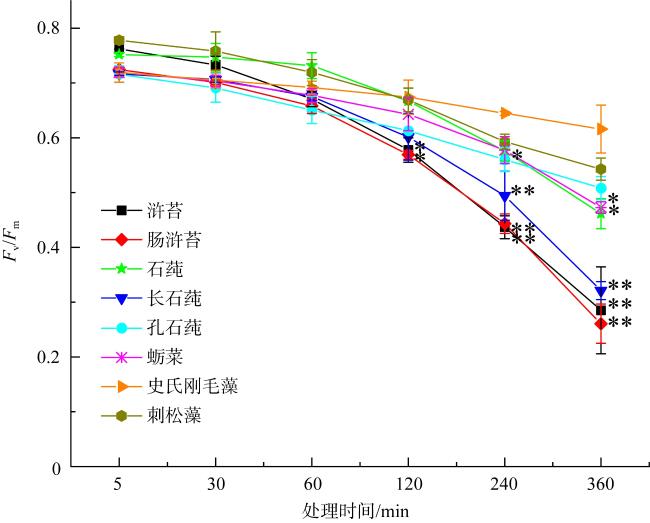
图2 不同处理时间的模拟酸雨(pH 4.5)胁迫对8种绿藻Fv/Fm的影响*和**表示在0.05和0.01水平上浒苔、肠浒苔、石莼、长石莼、孔石莼、蛎菜、刺松藻与史氏刚毛藻差异显著, 以史氏刚毛藻为对照 Fig. 2 Effects of simulated acid rain (pH 4.5) stress with different treatment times on Fv/Fm of eight species of green seaweeds. * and ** indicate that there are significant differences among U. prolifera, U. intestinalis, U. lactuca, U. stenophylla, U. pertusa, U. conglobata, C. fragile and C. stimpsonii at the level of 0.05 and 0.01. C. stimpsonii is control |
表1 叶绿素a含量(单位: mg·g-1)对模拟酸雨不同处理时间的胁迫响应Tab. 1 Stress response of chlorophyll a content (unit: mg·g-1) to different treatment times of simulated acid rain |
| 绿藻种类 | 0min | 30min | 60min | 120min | 240min | 360min |
|---|---|---|---|---|---|---|
| 浒苔 | 0.735±0.005 a | 0.728±0.010 a | 0.678±0.018 ab | 0.627±0.060 bc | 0.545±0.001 c | 0.446±0.010 d |
| 肠浒苔 | 0.479±0.005 a | 0.416±0.014 b | 0.401±0.017 bc | 0.370±0.007 bc | 0.356±0.022 cd | 0.312±0.021 d |
| 石莼 | 0.735±0.053 a | 0.733±0.021 a | 0.714±0.007 ab | 0.642±0.078 abc | 0.549±0.029 bc | 0.525±0.066 c |
| 长石莼 | 0.678±0.024 a | 0.674±0.039 a | 0.660±0.020 ab | 0.634±0.021 ab | 0.578±0.023 b | 0.479±0.034 c |
| 孔石莼 | 0.698±0.011 a | 0.702±0.032 a | 0.687±0.036 a | 0.664±0.012 a | 0.578±0.003 b | 0.514±0.011 b |
| 蛎菜 | 0.512±0.009 a | 0.477±0.010 ab | 0.444±0.034 ab | 0.420±0.012 ab | 0.410±0.008 ab | 0.366±0.093 b |
| 史氏刚毛藻 | 0.671±0.033 a | 0.672±0.012 a | 0.636±0.051 a | 0.616±0.009 a | 0.574±0.003 ab | 0.505±0.039 b |
| 刺松藻 | 0.675±0.009 a | 0.671±0.014 a | 0.660±0.018 a | 0.646±0.006 a | 0.586±0.016 b | 0.499±0.016 c |
注: 不同小写字母上标表示同一绿藻不同处理时间之间差异显著(p<0.05) |
表2 类胡萝卜素含量(单位: mg·g-1)对模拟酸雨不同处理时间的胁迫响应Tab. 2 Stress response of carotenoids content (unit: mg·g-1) to different treatment times of simulated acid rain |
| 绿藻种类 | 0min | 30min | 60min | 120min | 240min | 360min |
|---|---|---|---|---|---|---|
| 浒苔 | 0.444±0.017 a | 0.442±0.028 ab | 0.402±0.008 ab | 0.377±0.031 b | 0.303±0.021 c | 0.281±0.002 c |
| 肠浒苔 | 0.347±0.006 a | 0.313±0.008 ab | 0.305±0.012 b | 0.284±0.008 bc | 0.265±0.004 cd | 0.228±0.021 d |
| 石莼 | 0.470±0.033 a | 0.475±0.002 a | 0.450±0.019 a | 0.421±0.051 ab | 0.385±0.042 ab | 0.332±0.033 b |
| 长石莼 | 0.217±0.013 a | 0.212±0.003 ab | 0.188±0.012 bc | 0.171±0.006 cd | 0.155±0.003 d | 0.147±0.009 d |
| 孔石莼 | 0.567±0.031 a | 0.541±0.001 ab | 0.551±0.001 ab | 0.508±0.003 b | 0.448±0.001 c | 0.406±0.007 c |
| 蛎菜 | 0.284±0.017 a | 0.274±0.011 a | 0.270±0.001 a | 0.249±0.002 a | 0.233±0.014 a | 0.212±0.053 a |
| 史氏刚毛藻 | 0.396±0.041 a | 0.392±0.009 a | 0.388±0.008 ab | 0.372±0.017 ab | 0.365±0.022 ab | 0.313±0.026 b |
| 刺松藻 | 0.406±0.002 a | 0.400±0.005 a | 0.388±0.033 ab | 0.361±0.007 ab | 0.341±0.009 bc | 0.302±0.006 c |
注: 不同小写字母上标表示同一绿藻不同处理时间之间差异显著(p<0.05) |
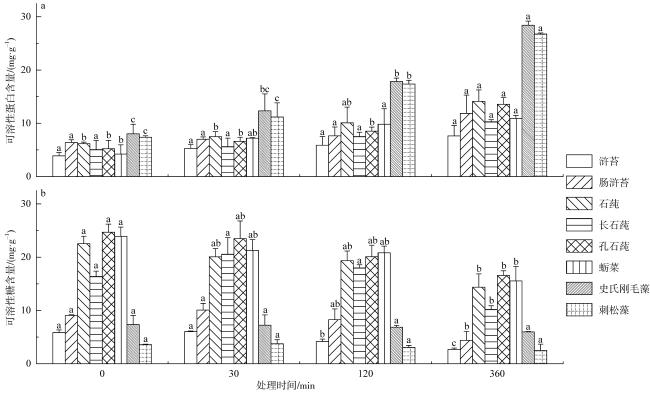
图3 可溶性蛋白含量(a)和可溶性糖含量(b)对模拟酸雨不同处理时间的胁迫响应不同小写字母表示同一绿藻不同处理时间之间差异显著(p<0.05) Fig. 3 Response of soluble protein content and soluble sugar content to simulated acid rain stress. Different lowercase letters indicate significant difference among different treatment times of the same green seaweed at 0.05 level |
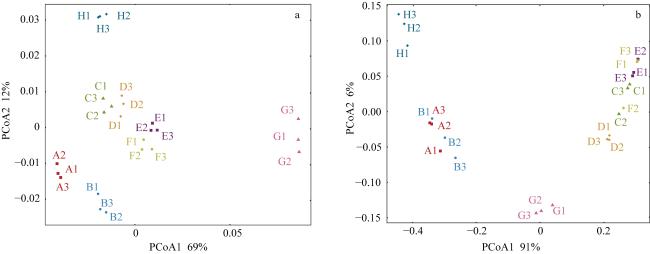
图5 绿藻Fv/Fm (a)和可溶性糖含量(b)的主坐标分析A1~A3: 浒苔; B1~B3: 肠浒苔; C1~C3: 石莼; D1~D3: 长石莼; E1~E3: 孔石莼; F1~F3: 蛎菜; G1~G3: 史氏刚毛藻; H1~H3: 刺松藻 Fig. 5 PcoA of Fv/Fm and soluble sugar content in green seaweeds. (a) PcoA of Fv/Fm in green seaweeds; (b) PcoA of soluble sugar content in green seaweeds. A1~A3: U. prolifera; B1~B3: U. intestinalis; C1~C3: U. lactuca; D1~D3: U. stenophylla; E1~E3: U. pertusa; F1~F3: U. congbata; G1~G3: C. stimpsonii; H1~H3: C. fragile |
| [1] |
卞雅姣, 黄洁, 孙其松, 等, 2013. 模拟酸雨对小麦产量及籽粒蛋白质和淀粉含量及组分的影响[J]. 生态学报, 33(15): 4623-4630.
|
| [2] |
丁兰平, 栾日孝, 2013. 中国海藻志: 第四卷绿藻门(第一册)[M]. 北京: 科学出版社: 44- 69, 135-137.
|
| [3] |
姜芳燕, 杨宁, 菅盼盼, 等, 2018. 基于 rbcL 和 tufA 基因的石莼和浒苔系统发育分析[J]. 基因组学与应用生物学, 37(7): 228-234.
|
| [4] |
李合生, 2000. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社: 182- 186, 194-203.
|
| [5] |
李信书, 徐军田, 何培民, 2011. 干燥和模拟酸雨胁迫对条斑紫菜光合作用的影响[J]. 水产科学, 30(5): 260-264.
|
| [6] |
连纲, 罗涛, 傅智慧, 等, 2021. 2001—2018 年浙江省酸雨变化特征及影响因素分析[J]. 中国环境监测, 37(4): 104-110.
|
| [7] |
廖源林, 蔡仕珍, 叶充, 等, 2015. 模拟酸雨对苦楝生理生态特性的影响[J]. 生态学杂志, 34(10): 2764-2770.
|
| [8] |
刘华, 吴国荣, 周耀民, 等, 2003. 模拟酸雨引起水体 pH 下降导致 Zn 对金鱼藻的毒害[J]. 环境科学学报, 23(4): 525-529.
|
| [9] |
宁波市生态环境局, 2022. 2021年宁波市生态环境状况公报[N]. 宁波日报, 2022-06-05(004).
NINGBO MUNICIPAL BUREAU OF ECOLOGY AND ENVIRONMENT, 2022. 2021 Ningbo ecological environment status bulletin[N]. Ningbo Daily, 2022-06-05(004) (in Chinese with English abstract).
|
| [10] |
齐泽民, 王玄德, 宋光煜, 2004. 酸雨对植物影响的研究进展[J]. 世界科技研究与发展, 26(2): 36-41.
|
| [11] |
水德聚, 王晓艳, 邵勤, 等, 2016. 模拟酸雨胁迫对油冬菜生理特性的影响[J]. 南方农业学报, 47(7): 1155-1158.
|
| [12] |
万文琴, 杜响, 秦欣, 等, 2022. 坛紫菜对干出和酸雨胁迫的生理响应[J]. 宁波大学学报(理工版), 35(2): 1-7.
|
| [13] |
王春燕, 2017. 不同模拟酸雨不同处理方式对华重楼生理特性的影响[D]. 成都: 四川师范大学.
|
| [14] |
王文彦, 2020. 模拟酸雨对玉米种子萌发和幼苗生长的影响[J]. 东北农业科学, 45(1): 21-24, 78.
|
| [15] |
王自发, 高超, 谢付莹, 2007. 中国酸雨模式研究回顾与所面临的挑战[J]. 自然杂志, 29(2): 78-82.
|
| [16] |
徐德才, 1995. 酸雨污染与防治: 浙江区域酸雨趋势与防治对策[J]. 能源环境保护, 9(4): 25-28.
|
| [17] |
徐晓婷, 李亚鹤, 王东, 等, 2016. 低盐胁迫下模拟酸雨对裂片石莼光合作用生理特性的影响[J]. 水产学报, 40(5): 731-739.
|
| [18] |
张银, 李文杰, 李玲丽, 等, 2020. 模拟酸雨对3种红豆杉幼苗生理指标的影响[J]. 西部林业科学, 49(1): 120-127.
|
| [19] |
张志良, 瞿伟菁, 2003. 植物生理学实验指导[M]. 第3版. 北京: 高等教育出版社: 121-127, 268-270.
|
| [20] |
赵会杰, 邹琦, 于振文, 2000. 叶绿素荧光分析技术及其在植物光合机理研究中的应用[J]. 河南农业大学学报, 34(3): 248-251.
|
| [21] |
周原也, 易晓芹, 周跃斌, 等, 2019. 模拟酸雨对茶树叶片光合色素含量及光合作用 CO2 响应的影响[J]. 分子植物育种, 17(21): 7201-7206.
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}