
含铁肠杆菌素受体调节蛋白VPA0148对副溶血弧菌毒力的影响
|
朱馨媛(1996—), 女, 江苏省南京市人, 硕士研究生, 从事副溶血弧菌毒力调控相关研究。email: |
Copy editor: 林强
收稿日期: 2020-12-14
修回日期: 2021-02-22
网络出版日期: 2021-02-25
基金资助
国家自然科学基金项目(31872597)
江苏省农业科技自主创新资金项目(CX[19]2033)
江苏省现代农业产业技术体系项目(JATS[2020]465)
版权
Effects of the ferric enterobactin receptor regulator VPA0148 on virulence of Vibrio parahaemolyticus
Copy editor: LIN Qiang
Received date: 2020-12-14
Revised date: 2021-02-22
Online published: 2021-02-25
Supported by
National Natural Science Foundation of China(31872597)
Jiangsu Agriculture Science and Technology Innovation Fund(CX[19]2033)
Earmarked Fund for Jiangsu Agricultural Industry Technology System(JATS[2020]465)
Copyright
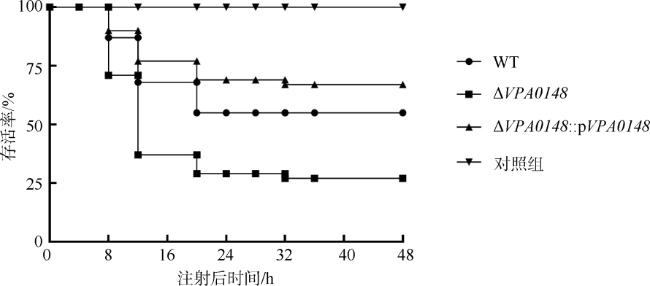
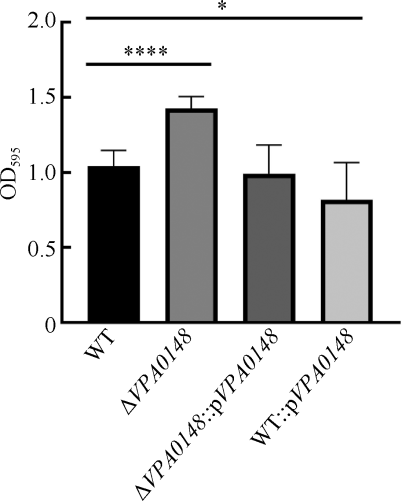
副溶血弧菌(Vibrio parahaemolyticus)是一种嗜盐性革兰氏阴性菌, 广泛分布于海洋环境中, 能够引起肠胃炎、伤口感染以及败血症, 是人类重要的病原菌之一。宿主体内被认为是一种低铁环境, 能够激发致细菌的毒力。副溶血弧菌能够分泌弧菌素或利用外源铁载体来获取铁。在低铁条件下, 编码双组份调控系统的基因簇VPA0148-VPA0149转录上调, 使含铁肠杆菌素受体PeuA转录出有活性的蛋白。VPA0148编码产物为一个响应调节因子, 具有一个磷酸基团接收结构域和一个DNA结合结构域。本研究发现, VPA0148的缺失增强了菌株的生物膜形成能力和对斑马鱼的致死能力, 同时能够促进菌株在低铁环境中的生长, 并增强菌株的群集运动。研究结果表明含铁肠杆菌素受体调节蛋白VPA0148对副溶血弧菌致病力具有调控作用, 增进了对铁调节副溶血弧菌毒力机制的认识。
关键词: 副溶血弧菌; 含铁肠杆菌素受体调节蛋白; 毒力; 运动能力; 生物膜
朱馨媛 , 刘敏 , 黄颖 , 赵哲 . 含铁肠杆菌素受体调节蛋白VPA0148对副溶血弧菌毒力的影响[J]. 热带海洋学报, 2021 , 40(6) : 93 -101 . DOI: 10.11978/2020146
Vibrio parahaemolyticus is a halophilic Gram-negative bacterium originally inhabiting marine environment. It is one of the primary human pathogenic bacteria, which causes gastroenteritis, wound infections and septicemia. The host environment is low-iron conditions, thus capable of inducing the virulence of pathogens. V. parahaemolyticus utilizes vibriocin or heterogenous siderophore to obtain iron. The transcription of gene cluster VPA0148-VPA0149 (encoding a two-component regulatory system) is upregulated under iron-limiting conditions, which increases the active transcriptional form of ferric enterobactin receptor gene peuA. VPA0148 encodes a response regulator containing a phosphoacceptor receiver (REC) domain and a DNA-binding domain. In this study, the deletion of VPA0148 enhanced the lethality to zebrafish as well as biofilm formation in V. parahaemolyticus. Moreover, under iron-limiting conditions, the growth and swarming motility were improved in VPA0148 deletion mutant. The results indicate that VPA0148 contributes to regulating the pathogenesis of V. parahaemolyticus. This study provides an important insight for further understanding the mechanism of how iron regulates virulence of V. parahaemolyticus.
Key words: Vibrio parahaemolyticus; VPA0148; virulence; swarming; biofilm
表1 本研究中使用的菌株和质粒Tab. 1 Strains and plasmids |
| 名称 | 描述 | 来源或参考文献 | |
|---|---|---|---|
| 菌株 | RIMD2210633 (WT) | 副溶血弧菌血清型O3:K6临床分离株; 野生型菌株 | (Makino et al, 2003) |
| ΔVPA0148 | 副溶血弧菌VPA0148基因缺失株 | 本研究 | |
| ΔVPA0148::pVPA0148 | 副溶血弧菌VPA0148基因回补株 | 本研究 | |
| WT::pVPA0148 | 含有VPA0148-pBBR1-MCS-1质粒的副溶血弧菌野生型菌株 | 本研究 | |
| S17-1λpir | Thi pro hsdR hsdM+ recA RP4-2-Tc::Mu-Km::Tn7λpir接合供体菌, 大肠杆菌 | (Milton et al, 1992) | |
| 质粒 | pDM4 | Cmr, 含依赖π蛋白oriP6K复制子和sacBR基因的自杀质粒, | (Milton et al, 1996) |
| pBBR1-MCS-1 | Cmr, 广宿主蛋白表达或克隆载体质粒, 高拷贝 | (Kovach et al, 1994) | |
| VPA0148-pBBR1-MCS-1 | Cmr, 插入VPA0148基因及其启动子片段的pBBR1-MCS-1质粒 | 本研究 | |
| ΔVPA0148-pDM4 | Cmr, 插入VPA0148基因缺失片段的pDM4质粒 | 本研究 |
表2 本研究中所使用的引物Tab. 2 Primers in this research |
| 名称 | 序列(5′—3′) | 用途 |
|---|---|---|
| d0148-1F | CCCTCGAGGTTGCCAGTTGGCACGATG | 用于扩增VPA0148侧翼区域进行基因敲除 |
| d0148-1R | CCACAGATTCTAAGCCAAATTACCACTTAAATAAAAATGG | |
| d0148-2F | TATTTAAGTGGTAATTTGGCTTAGAATCTGTGGGCGCGTG | |
| d0148-2R | CGAGCTCTCATCAATGCTGCGTTTGAATTG | |
| p0148-F | GTCGACGGTATCGATAAGCTTGTTGCCAGTTGGCACGATG | 扩增VPA0148启动子及基因片段进行回补质粒构建 |
| p0148-R | GGGCGAATTGGAGCTTCACTTGTCATCGTCATCCTTGTAATCCGCGCCCACAGATTCTA | |
| RT0148F | CATTTGTAATACCGCCCAGAG | 荧光定量 |
| RT0148R | GATCACCACGCCTTGATTCTT |
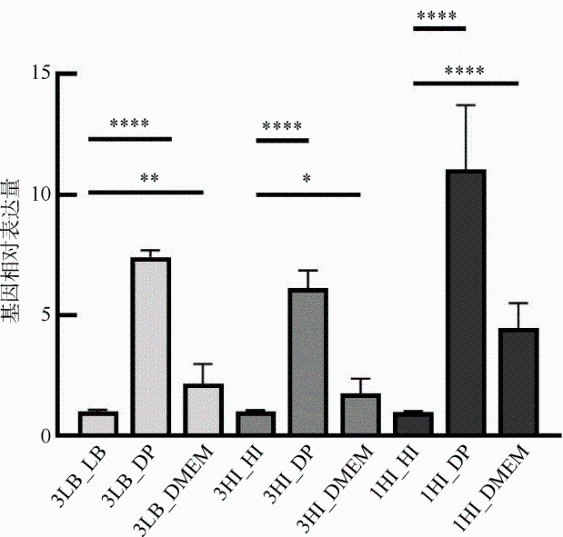
图2 相同NaCl浓度和限铁条件下VPA0148的转录分析3LB_LB: 含3% NaCl的LB培养基中培养的菌株转接到含3% NaCl的LB培养基中; 3LB_DP: 含3% NaCl的LB培养基中培养的菌株转接到含3% NaCl的添加了螯合剂的LB培养基中; 3LB_DMEM: 含3% NaCl的LB培养基中培养的菌株转接到DMEM培养基中; 3HI_HI: 含3% NaCl的心浸液培养基中培养的菌株转接到含3% NaCl的心浸液培养基中; 3HI_DP: 含3% NaCl的心浸液培养基中培养的菌株转接到含3% NaCl的添加了螯合剂的心浸液培养基中; 3HI_DMEM: 含3% NaCl的心浸液培养基中培养的菌株转接到DMEM培养基中; 1HI_HI: 含1% NaCl的普通心浸液培养基中培养的菌株转接到含1% NaCl的心浸液培养基中; 1HI_DP: 含1% NaCl的普通心浸液培养基中培养的菌株转接到含1% NaCl的添加了螯合剂的心浸液培养基; 1HI_DMEM: 含1% NaCl的普通心浸液培养基中培养的菌株转接到DMEM培养基中。qPCR检测VPA0148的转录水平。显著性: *P<0.05, **P<0.01, ****P<0.0001 Fig. 2 Relative VPA0148 mRNA transcriptional levels under different concentrations of NaCl and iron-limiting condition |
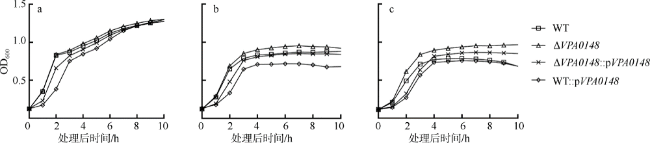
图3 VPA0148对副溶血弧菌在低铁条件下生长的影响本实验使用的心浸液培养基中含1% NaCl。将过夜培养的菌株分别转接到心浸液培养基(a)或含螯合剂的心浸液培养基(b)中; 将在含螯合剂的心浸液培养基中培养的菌株, 转接到含螯合剂的心浸液培养基(c)中, 测定生长曲线。WT: 野生型菌株; ΔVPA0148: VPA0148缺失株; ΔVPA0148::pVPA0148: VPA0148缺失株中回补带有VPA0148基因的pBBR1-MCS-1质粒; WT::pVPA0148: 野生型菌株中回补带有VPA0148基因的pBBR1-MCS-1质粒 Fig. 3 VPA0148 inhibiting the growth of V. parahaemolyticus under low-iron condition |
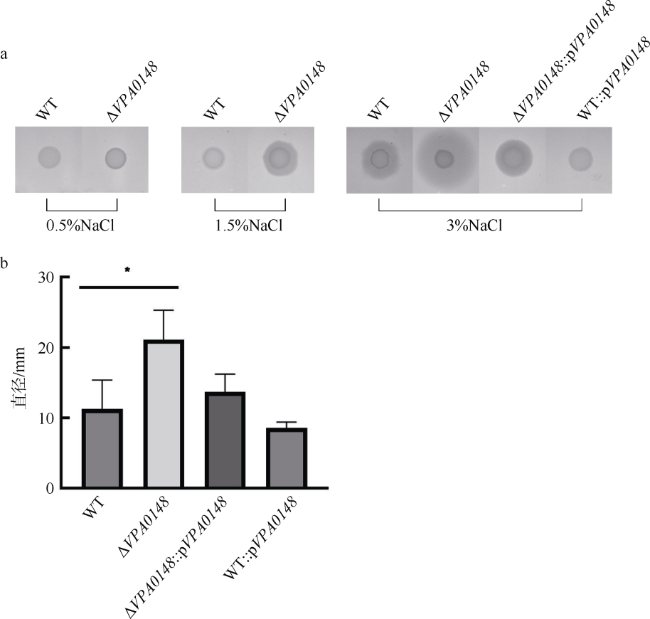
图4 VPA0148对副溶血弧菌群集运动活性的影响a. 不同NaCl浓度心浸液-琼脂平板上菌株的运动范围; b. 3% NaCl浓度心浸液-琼脂平板上不同菌株的运动范围。WT: 野生型菌株; ΔVPA0148: VPA0148缺失株; ΔVPA0148::pVPA0148: VPA0148缺失株中回补带有VPA0148基因的pBBR1-MCS-1质粒; WT::pVPA0148: 野生型菌株中回补带有VPA0148基因的pBBR1-MCS-1质粒。显著性: *P<0.05 Fig. 4 VPA0148 inhibiting the swarming activity of V. parahaemolyticus |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}