
广西北海红树林宜林滩涂大型底栖动物群落结构特征
|
耿婉璐(1997—), 女, 河南省许昌市人, 硕士研究生, 主要从事湿地生态修复研究。email: |
Copy editor: 孙翠慈
收稿日期: 2023-04-12
修回日期: 2023-05-25
网络出版日期: 2023-06-28
基金资助
国家自然科学基金项目(42141016)
自然资源部第四海洋研究所基本科研业务费专项资金资助项目(202005)
Structural characteristics of macrobenthic communities at intertidal zone for mangrove in Beihai, Guangxi
Copy editor: SUN Cuici
Received date: 2023-04-12
Revised date: 2023-05-25
Online published: 2023-06-28
Supported by
National Natural Science Foundation of China(42141016)
Scientific Research Fund of the Fourth Institute of Oceanography,MNR(202005)
耿婉璐 , 邢永泽 , 张秋丰 , 管卫兵 . 广西北海红树林宜林滩涂大型底栖动物群落结构特征[J]. 热带海洋学报, 2024 , 43(1) : 107 -115 . DOI: 10.11978/2023047
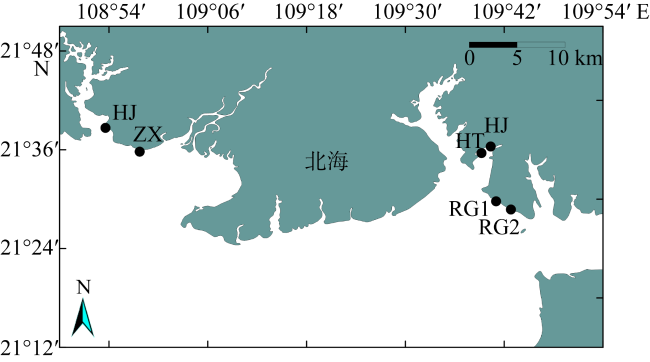
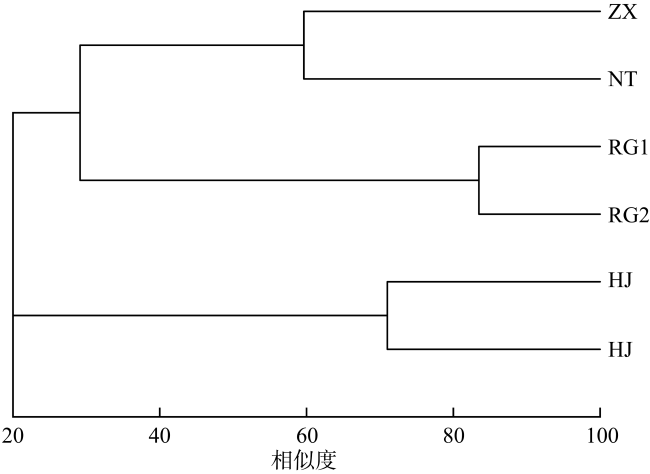
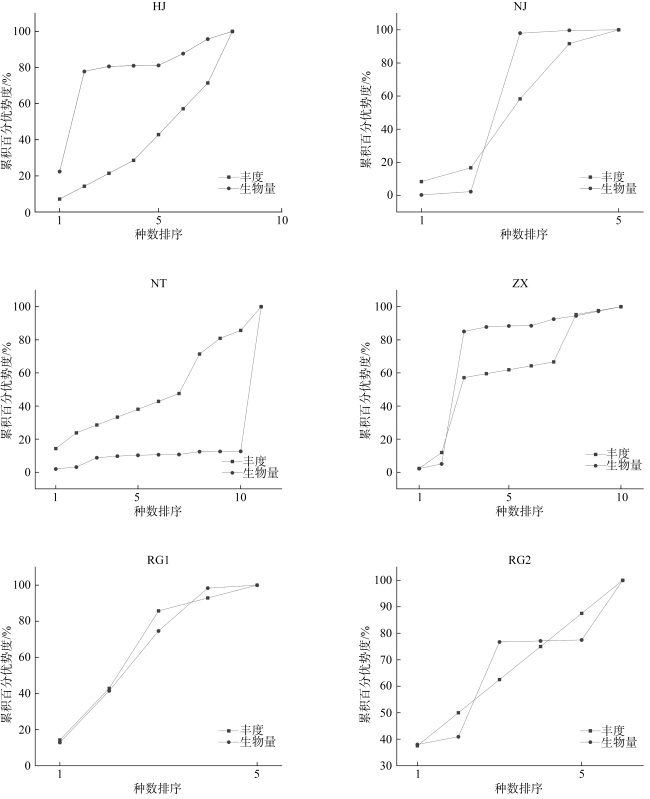
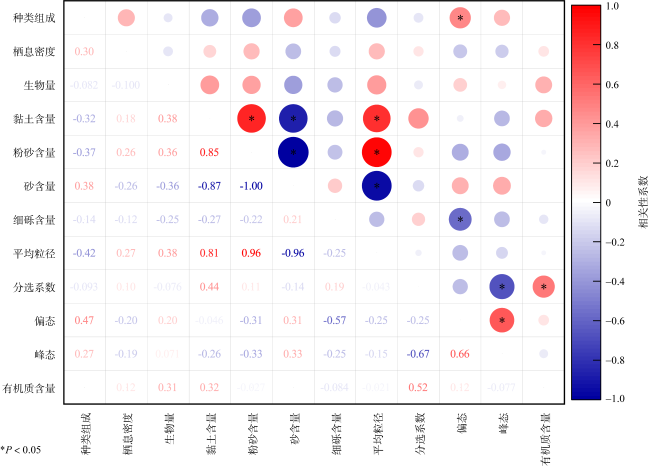
In order to understand the characteristics of macrobenthic communities in the intertidal zone for mangroves, two sections of bare flat, two sections with a small number of Spartina alterniflora distribution and two sections covered by Spartina alterniflora were selected in Beihai for ecological investigation, and the community structure of macrobenthic animals and its relationship with environmental factors were analyzed. There were 29 macrobenthic species in the survey area, and there were significant differences in mollusc biomass, arthropod habitat density and crab burrow number in different habitats (P< 0.05), and the species composition of macrobenthic animals was positively correlated with sediment skewness (P< 0.05). The HJ community structure was stable in the bare flat, but the community structure of macrobenthic animals in the other plots was unstable. The growth of Spartina alterniflora affects the structure of macrobenthic communities.
Key words: macrobenthic animals; community structure; Spartina alterniflora; mangrove
表1 不同样地土壤理化因子Tab.1 Soil physical and chemical factors in different places |
| 样地类型 | 样地名称 | 有机质/% | 黏土/% | 粉砂/% | 砂/% | 细砾/% |
|---|---|---|---|---|---|---|
| 光滩 | HJ | 0.47±0.05a | 1.42±0.21ab | 28.56±3.48bc | 69.83±3.60ab | 0.19±0.11 |
| NJ | 0.47±0.03a | 2.41±0.21b | 37.91±3.90c | 59.68±4.10a | - | |
| 平均值 | 0.48±0.09 | 1.92±0.01* | 33.23±0.15* | 64.76±0.18 | 0.10±0.039 | |
| 少量互花米草 | ZX | 1.88±0.12ab | 2.18±0.09b | 29.95±0.64bc | 67.56±0.64ab | 0.31±0.18 |
| NT | 3.08±0.45b | 1.97±0.06ab | 26.21±0.63abc | 71.82±0.61abc | - | |
| 平均值 | 2.48±0.12* | 2.08±0.01b | 28.08±0.01 | 69.69±0.01 | 0.16±0.06 | |
| 互花米草 | RG1 | 0.77±0.22a | 1.23±0.36a | 17.88±1.71ab | 80.75±2.02bc | 0.14±0.08 |
| RG2 | 1.35±0.33a | 0.90±0.15a | 15.19±0.29a | 83.83±0.37c | 0.08±0.04 | |
| 平均值 | 1.06±0.38 | 1.07±0.08 | 16.54±0.50 | 82.29±0.59* | 0.11±0.01 |
注: a, b, c表示不同调查站位之间差异显著(P< 0.05); *表示各样地类型之间差异显著(P< 0.05); -表示调查站位该因子含量为0 |
表2 不同样地大型底栖动物物种数Tab. 2 The number of macrobenthic species in different places |
| 样地类型 | 样地名称 | 物种数 | ||||||
|---|---|---|---|---|---|---|---|---|
| 软体动物 | 节肢动物 | 纽形动物 | 星虫动物 | 环节动物 | 脊索动物 | 总计 | ||
| 光滩 | HJ | 7 | 0 | 1 | 0 | 0 | 0 | 8 |
| NJ | 3 | 1 | 0 | 0 | 1 | 0 | 5 | |
| 合计 | 10 | 1 | 1 | 0 | 1 | 0 | 13 | |
| 少量 互花米草 | ZX | 7 | 1 | 0 | 0 | 0 | 1 | 9 |
| NT | 3 | 5 | 0 | 1 | 1 | 0 | 10 | |
| 合计 | 10 | 6 | 0 | 1 | 1 | 1 | 19 | |
| 互花米草 | RG1 | 4 | 1 | 0 | 0 | 0 | 0 | 5 |
| RG2 | 1 | 4 | 0 | 0 | 1 | 0 | 6 | |
| 合计 | 5 | 5 | 0 | 0 | 1 | 0 | 11 | |
表3 不同样地大型底栖动物优势种Tab. 3 Dominant species of macrofauna in different places |
| 样地类型 | 优势种 | 优势度指数(Y) |
|---|---|---|
| 光滩 | 中国绿螂 Glauconome chinensis | 0.095 |
| 红树蚬 Geloina coaxans | 0.037 | |
| 少量互花米草 | 珠带拟蟹守螺 Cerithidea cingulata | 0.091 |
| 红果滨螺 Littorina coccinea | 0.036 | |
| 互花米草 | 凹指招潮 Uca vocans | 0.074 |
| 中国绿螂 Glauconome chinensis | 0.033 |
表4 不同样地大型底栖动物生物量Tab. 4 Biomass of macrofauna in different places |
| 样地类型 | 样地名称 | 生物量/(g·m−2) | |||
|---|---|---|---|---|---|
| 软体动物 | 节肢动物 | 其他动物 | 合计 | ||
| 光滩 | HJ | 182.83 | 0 | 0.64 | 183.47 |
| NJ | 176.16 | 2.45 | 2.35 | 180.96 | |
| 平均值 | 179.49±3.34a | 1.23±1.23a | 1.49±0.86a | 182.23±1.26a | |
| 少量互花米草 | ZX | 318.51 | 10.19 | 5.6 | 334.29 |
| NT | 793.07 | 38.61 | 19.68 | 851.36 | |
| 平均值 | 555.79±237.28b | 24.4±14.21a | 12.64±7.04a | 592.83±258.54b | |
| 互花米草 | RG1 | 693.87 | 66.83 | 0 | 760.69 |
| RG2 | 703.95 | 3.79 | 5.49 | 713.23 | |
| 平均值 | 698.91±5.04b | 35.31±31.52a | 2.75±2.75a | 736.96±23.73b | |
注: a, b表示不同样地类型之间差异显著(P< 0.05) |
表5 不同样地大型底栖动物栖息密度Tab. 5 Density of macrofauna in different places |
| 样地类型 | 样地名称 | 栖息密度/(ind.·m−2) | ||||
|---|---|---|---|---|---|---|
| 软体动物 | 节肢动物 | 其他动物 | 合计 | 蟹洞 | ||
| 光滩 | HJ | 70 | 0 | 6 | 76 | 304 |
| NJ | 54 | 6 | 6 | 66 | 1008 | |
| 平均值 | 62±8a | 3±3a | 6a | 71±5a | 656±352a | |
| 少量互花米草 | ZX | 214 | 6 | 6 | 226 | 1158 |
| NT | 32 | 27 | 54 | 113 | 2603 | |
| 平均值 | 123±91a | 17±11ab | 30±24a | 169±57b | 1880±723b | |
| 互花米草 | RG1 | 43 | 32 | 0 | 75 | 587 |
| RG2 | 6 | 22 | 16 | 44 | 358 | |
| 平均值 | 25±19a | 27±5b | 8±8a | 59±16a | 472±115a | |
注: a, b表示不同样地类型之间差异显著(P< 0.05) |
表6 不同样地大型底栖动物群落的物种多样性指数Tab. 6 Species diversity index for macrobenthic communities in different places |
| 样地类型 | 样地名称 | 多样性指数(H′) | 丰富度指数(d) | 均匀度指数(J) |
|---|---|---|---|---|
| 光滩 | HJ | 2.11 | 1.12 | 0.70 |
| NJ | 1.63 | 0.66 | 0.70 | |
| 平均值 | 1.87a | 0.89ab | 0.70a | |
| 少量互花米草 | ZX | 1.66 | 1.02 | 0.52 |
| NT | 2.22 | 1.32 | 0.67 | |
| 平均值 | 1.94a | 1.17a | 0.59b | |
| 互花米草 | RG1 | 1.38 | 0.64 | 0.59 |
| RG2 | 1.67 | 0.92 | 0.65 | |
| 平均值 | 1.52a | 0.78b | 0.62ab |
注: a, b表示各样地类型之间差异显著(P< 0.05) |
| [1] |
鲍士旦, 2000. 土壤农化分析(第三版)[M]. 北京: 中国农业出版社: 25-38.
|
| [2] |
蔡永久, 龚志军, 秦伯强, 2010. 太湖大型底栖动物群落结构及多样性[J]. 生物多样性, 18(1): 50-59.
|
| [3] |
冯建祥, 黄茜, 陈卉, 等, 2018. 互花米草入侵对盐沼和红树林滨海湿地底栖动物群落的影响[J]. 生态学杂志, 37(3): 943-951.
|
| [4] |
黄雅琴, 王建军, 何雪宝, 等, 2020. 三沙湾互花米草(Spartina alterniflora) 入侵对大型底栖动物群落结构的影响[J]. 海洋与湖沼, 51(3): 506-519.
|
| [5] |
纪莹璐, 蒲思潮, 陶卉卉, 等, 2022. 丁字湾盐沼湿地不同植被生境大型底栖动物群落结构研究[J]. 山东农业大学学报(自然科学版), 53(3): 412-420.
|
| [6] |
赖廷和, 何斌源, 黄中坚, 等, 2019. 防城河口湾潮间带大型底栖动物群落结构研究[J]. 热带海洋学报, 38(2): 67-77.
|
| [7] |
刘士龙, 秦旭东, 王广军, 等, 2019. 2017 年夏季北海市冯家江入海口红树林潮间带大型底栖动物群落结构及多样性[J]. 湿地科学, 17(3): 352-358.
|
| [8] |
陆琳莹, 邵学新, 杨慧, 等, 2020. 浙江滨海湿地互花米草生长性状对土壤化学因子的响应[J]. 林业科学研究, 33(5): 177-183.
|
| [9] |
马克平, 1994. 生物群落多样性的测度方法Ⅰα多样性的测度方法(上)[J]. 生物多样性, 1994(3): 162-168.
|
| [10] |
马文刚, 夏景全, 魏一凡, 等, 2022. 三亚蜈支洲岛海洋牧场近岛区底表大型底栖动物群落结构及评价[J]. 热带海洋学报, 41(3): 135-146.
|
| [11] |
仇乐, 刘金娥, 陈建琴, 等, 2010. 互花米草扩张对江苏海滨湿地大型底栖动物的影响[J]. 海洋科学, 34(8): 50-55.
|
| [12] |
任海庆, 袁兴中, 刘红, 等, 2015. 环境因子对河流底栖无脊椎动物群落结构的影响[J]. 生态学报, 35(10): 3148-3156.
|
| [13] |
沈永明, 王艳芳, 陈寿军, 等, 2013. 互花米草盐沼湿地大型底栖动物时空分布特征[J]. 地理研究, 32(4): 638-644.
|
| [14] |
谢志发, 何文珊, 刘文亮, 等, 2008. 不同发育时间的互花米草盐沼对大型底栖动物群落的影响[J]. 生态学杂志, 27(1): 63-67.
|
| [15] |
杨泽华, 童春富, 陆健健, 2007. 盐沼植物对大型底栖动物群落的影响[J]. 生态学报, 27(11): 4387-4393.
|
| [16] |
袁涛萍, 李恒翔, 李路, 等, 2017. 夏季大亚湾大型底栖动物群落结构[J]. 热带海洋学报, 36(1): 41-47.
|
| [17] |
赵彩云, 李俊生, 宫璐, 等, 2014. 广西北海市滨海湿地互花米草入侵对大型底栖动物的影响[J]. 湿地科学, 12(6): 733-739.
|
| [18] |
赵永强, 曾江宁, 陈全震, 等, 2009. 不同互花米草(Spartina alterniflora Loisel)密度生境中大型底栖动物群落格局[J]. 自然资源学报, 24(4): 630-639.
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}