
大亚湾夏季浮游群落生产代谢特征及其影响因素*
|
李尧(1996—), 女, 湖北省荆州市人, 硕士研究生, 从事海洋生态学研究。email: |
Copy editor: 林强
收稿日期: 2020-12-24
修回日期: 2021-03-04
网络出版日期: 2021-03-04
基金资助
国家自然科学基金(41890853)
国家重点研发计划(2017YFC0506302)
国家重点研发计划(2016YFC0502805)
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0404)
国家科技基础资源调查专项(2018FY10010005)
国家重点基础研究发展计划(2015CB452904)
版权
Production and metabolism characteristics of planktonic community and their influencing factors in Daya Bay during summer*
Copy editor: LIN Qiang
Received date: 2020-12-24
Revised date: 2021-03-04
Online published: 2021-03-04
Supported by
National Natural Science Foundation(41890853)
National Key Research and Development Program of China(2017YFC0506302)
National Key Research and Development Program of China(2016YFC0502805)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory(Guangzhou)(GML2019ZD0404)
National Science and Technology Fundamental Resources Investigation Program of China(2018FY10010005)
Key Project of Chinese National Programs for Fundamental Research and Development(2015CB452904)
Copyright
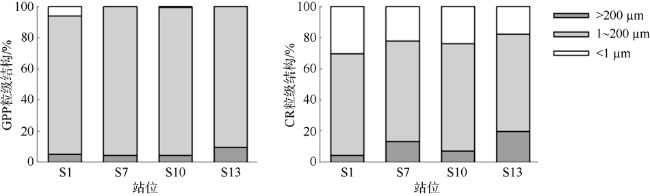
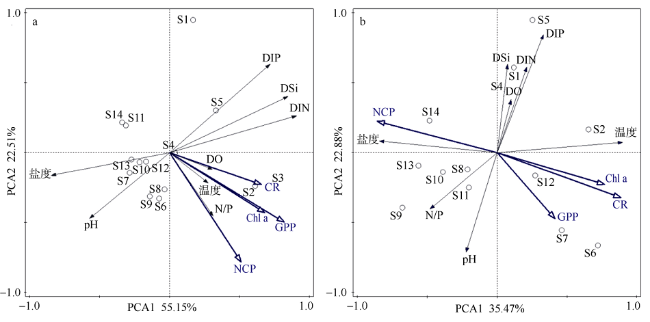
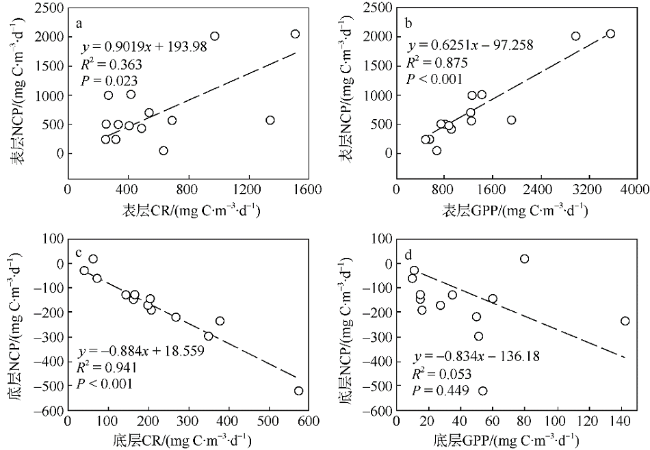
2017年8月对大亚湾海域浮游群落初级生产、群落呼吸代谢及其平衡特征进行了研究, 并分析了其潜在环境影响因素以及对沿岸生态系统功能与健康的指示作用。研究结果表明, 大亚湾夏季海水表层呈自养状态, 而底层呈异养状态, 群落总初级生产力(gross primary productivity, GPP)、呼吸代谢速率(community respiration, CR)与净生产力(net commutnitiy production, NCP)在表层分别为1335.36±910.12、597.86±403.30和737.50±608.22mg C·m-3·d-1; 在底层分别为43.65±37.05、216.25±147.28和-160.27±137.01mg C·m-3·d-1。海湾整体呈自养状态, 水柱平均NCP为233.41±248.88mg C·m-3·d-1; 部分沿岸水域存在异养状态。1~200μm粒级浮游生物是GPP和CR的主要贡献者。相关性分析和主成分分析显示, NCP在表层受GPP和CR共同调控, 且与浮游植物生物量和营养盐正相关; 而在底层主要受CR影响, 且与盐度正相关。大亚湾夏季群落生产代谢平衡存在明显的水平和垂向变化, NCP的区域差异与潜在波动性对该海湾生态系统稳定性及健康状况有重要的指示作用。
李尧 , 向晨晖 , 江志坚 , 宋星宇 . 大亚湾夏季浮游群落生产代谢特征及其影响因素*[J]. 热带海洋学报, 2021 , 40(6) : 83 -92 . DOI: 10.11978/2020149
In August 2017, the primary production, community respiration and their metabolic balance status of the planktonic community in Daya Bay were studied, and the potential environmental impact factors and their indicator effects on the function and health of the offshore ecosystem were analyzed. Our results showed that Daya Bay was autotrophic in surface water but heterotrophic in the bottom layer. The gross primary productivity (GPP), community respiration (CR) and net community production (NCP) were 1335±910.12, 597.86±403.30 and 737.50±608.22 mg C·m-3·d-1 in surface water and 43.65±37.05, 216.25±147.28 and -160.27±137.01 mg C·m-3·d-1 in the bottom, respectively. Overall, Daya Bay was autotrophic with an average NCP of 233.41±248.88 mg C·m-3·d-1 in the water column. Size-fractioned result of plankton community metabolism showed that plankton with the cell size of 1-200 μm was the main contributor to GPP and CR in surface water. Correlation analysis and principal component analysis showed that NCP was jointly regulated by GPP and CR in surface water and was positively correlated with phytoplankton biomass as well as nutrient concentration; in the bottom, it was mainly affected by CR and was positively correlated with salinity. The planktonic metabolism balance in Daya Bay during summer varied horizontally and vertically. The regional differences and potential fluctuations of NCP are important indicators of ecosystem stability and health in the bay.
Key words: Primary production; Community respiration; Metabolic balance; Daya Bay
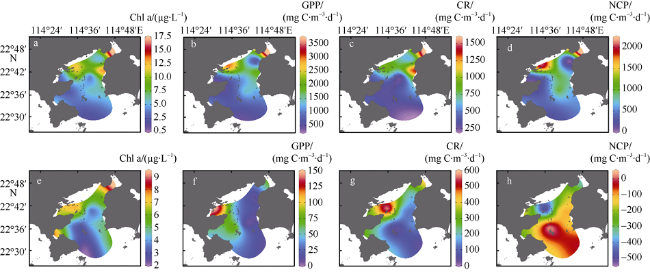
图2 大亚湾夏季表层(a—d)和底层(e—h)叶绿素a(Chl a)、群落总初级生产力(GPP)、群落呼吸速率(CR)和群落净生产力(NCP)的分布特征Fig. 2 Distribution of Chl a, gross primary productivity (GPP), community respiration (CR), and net community productivity (NCP) in surface water (a, b, c, d) and bottom water (e, f, g, h) of Daya Bay in summer |
表1 大亚湾夏季海水理化因子分布特征Tab. 1 Environmental parameters of Daya Bay in summer |
| 理化因子 | 表层 | 底层 | ||
|---|---|---|---|---|
| 范围 | 均值±方差 | 范围 | 均值±方差 | |
| pH | 7.84~8.49 | 8.18±0.18 | 7.75~8.35 | 8.05±0.17 |
| DO/(mg·L-1) | 7.15~12.72 | 9.75±1.77 | 5.03~7.72 | 6.38±0.83 |
| 盐度/‰ | 15.73~32.16 | 27.1±5.13 | 31.5~34.04 | 32.96±0.95 |
| 温度/℃ | 28.3~30.6 | 29.78±0.79 | 25.9~28.7 | 27.57±0.89 |
| PO3- 4/(μmol·L-1) | 0.04~1.49 | 0.39±0.51 | 0.04~0.54 | 0.2±0.14 |
| SiO2- 3/(μmol·L-1) | 1.03~31.86 | 11.16±11.81 | 3.67~23.92 | 10.71±6.51 |
| DIN/(μmol·L-1) | 1.75~85.1 | 14.12±23.03 | 2.78~12.27 | 4.72±2.41 |
| N/P | 6.35~116.31 | 39.89±31.25 | 11.16~106.95 | 31.81±24.5 |
表2 大亚湾夏季生产代谢特征与环境参数的相关性Tab. 2 Correlation between planktonic metabolism with environmental parameters of Daya Bay in summer |
| 环境参数 | 相关性指数 | |||||
|---|---|---|---|---|---|---|
| 表层NCP | 表层CR | 表层GPP | 底层NCP | 底层CR | 底层GPP | |
| Chl a | 0.712** | 0.96* | 0.901** | -0.678* | 0.672* | 0.216 |
| pH | -0.338 | -0.379 | -0.394 | -0.127 | 0.127 | 0.043 |
| DO | 0.279 | 0.369 | 0.35 | -0.354 | 0.222 | -0.402 |
| 盐度 | -0.667** | -0.417 | -0.631* | 0.694** | -0.736** | -0.41 |
| 温度 | 0.241 | 0.292 | 0.29 | -0.633* | 0.661* | 0.332 |
| PO3- 4 | 0.216 | 0.171 | 0.22 | -0.062 | 0.009 | -0.189 |
| SiO2- 3 | 0.587* | 0.539* | 0.631* | -0.021 | -0.075 | -0.373 |
| DIN | 0.678** | 0.381 | 0.622* | 0.061 | -0.081 | -0.099 |
注: *表示显著相关(P<0.05); **表示显著相关(P<0.01) |
| [1] |
陈得仿, 2019. 大亚湾渔业资源概况及黑鲷的生长、繁殖研究[D]. 上海: 上海海洋大学.
|
| [2] |
黄小平, 黄良民, 宋金明, 等, 2019. 营养物质对海湾生态环境影响的过程与机理[M]. 北京: 科学出版社: 16-17.
|
| [3] |
林晓娟, 高姗, 仉天宇, 等, 2018. 海水富营养化评价方法的研究进展与应用现状[J]. 地球科学进展, 33(4):373-384.
|
| [4] |
宋星宇, 黄良民, 张建林, 等, 2004. 大鹏澳浮游植物现存量和初级生产力及N:P值对其生长的影响[J]. 热带海洋学报, 23(5):34-41.
|
| [5] |
王娜, 林伟, 陈炳章, 等, 2014. 南海北部及台湾海峡夏季自养与异养区域的分布[J]. 热带海洋学报, 33(4):61-68.
|
| [6] |
王友绍, 王肇鼎, 黄良民, 2004. 近20年来大亚湾生态环境的变化及其发展趋势[J]. 热带海洋学报, 23(5):85-95.
|
| [7] |
谢福武, 宋星宇, 谭烨辉, 等, 2019. 模拟升温和营养盐加富对大亚湾浮游生物群落代谢的影响[J]. 热带海洋学报, 38(2):48-57.
|
| [8] |
徐恭昭, 1989. 大亚湾环境与资源[M]. 合肥: 安徽科学技术出版社: 5-9.
|
| [9] |
曾阳, 黄毅彬, 黄邦钦, 2015. 南黄海大型水母暴发区浮游生物群落代谢及细菌生产力[J]. 应用海洋学学报, 34(4):542-548.
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}