
广西北部湾海域织锦巴非蛤精巢发育、精子发生及超微结构研究
|
徐炳杰(1997—), 男, 广西梧州市人, 硕士研究生, 从事海洋贝类遗传与育种研究。email: |
Copy editor: 殷波
收稿日期: 2023-06-30
修回日期: 2023-08-22
网络出版日期: 2023-08-29
基金资助
广西创新驱动发展专项(Guike AA19254032)
Spermary development, spermatogenesis and sperm ultrastructure of Paphia textile in the Beibu Gulf, Guangxi
Copy editor: YIN Bo
Received date: 2023-06-30
Revised date: 2023-08-22
Online published: 2023-08-29
Supported by
Guangxi Innovation Driven Development Project(Guike AA19254032)
为了解广西北部湾海域织锦巴非蛤(Paphia textile)精巢发育、精子发生及成熟精子的超微结构, 文章采用解剖观察、精巢组织切片、扫描和透射电镜技术进行了研究。结果表明, 所采集织锦巴非蛤中未发现雌雄同体, 观察的个体皆为雌雄异体, 将其精巢发育划分为增殖期、生长期、成熟期、排放期、休止期5个时期, 织锦巴非蛤精巢成熟与排放期在9 月—翌年1月。精子发生分为精原细胞期、初级精母细胞期、次级精母细胞期、精细胞期、成熟精子期5个时期。织锦巴非蛤成熟精子由头部、中部和尾部组成, 为典型的鞭毛型精子, 全长约为46.04μm。精子头部分为顶体和精核, 顶体为圆锥形, 精核呈长圆柱形, 前端直径约为0.71μm, 后端直径约为1.34μm; 中部由4个环绕成瓣状的线粒体与2个相互垂直的近、远端中心粒组成; 尾部鞭毛长约38.8μm, 直径约为0.21μm, 外被质膜, 为典型“9+2”双联体微管结构。研究结果为织锦巴非蛤的繁殖生物学、人工苗种繁育及种质资源保护提供研究基础。
徐炳杰 , 刘一鸣 , 邢清淦 , 连昌朋 , 吴韬 , 潘英 . 广西北部湾海域织锦巴非蛤精巢发育、精子发生及超微结构研究[J]. 热带海洋学报, 2024 , 43(2) : 59 -68 . DOI: 10.11978/2023090
In this study, anatomical observation, spermary tissue section, scanning and transmission electron microscopy techniques were used to study the spermary development, spermatogenesis and ultrastructure of mature sperm of Paphia textile in Beibu Gulf of Guangxi. The results showed that no hermaphrodites were found in the collected samples, and all the observed individuals were dioecious. The spermary development of P. textile was divided into five main stages: proliferating stage, growing stage, maturing stage, spawning stage, and resting stage. The maturing stage and spawning stage of P. textile was from September to January of the following year. Sperm development is divided into five stages: spermatogonia stage, primary spermatocyte stage, secondary spermatocyte stage, spermatid stage and mature sperm stage. The mature sperm of P. textile consisting of the head, middle and tail, overall length approximately 46.04 μm. The sperm head is divided into the acrosome and the nucleus. The acrosome is conical in shape and the nucleus is long and cylindrical. The anterior end was 0.71 μm in diameter and the posterior end was about 1.34 μm in diameter. The middle part consisted of four mitochondria encircled in a circular shape and two mutually perpendicular proximal and distal centrioles. The tail flagellum was about 38.8 μm long and 0.21 μm in diameter, and the external plasma membrane was covered with a typical “9 + 2” duplex microtubule structure. The results of this study provide basic research information for the reproductive biology, artificial seed breeding and germplasm resource conservation of P. textile.
表1 织锦巴非蛤形态参数Tab.1 Morphological parameters of P. textile |
| 参数 | 壳长/mm | 壳宽 /mm | 壳高/mm | 体质量/g | 软体质量/g |
|---|---|---|---|---|---|
| 平均值±标准差 | 62.73±5.39 | 23.45±2.62 | 36.90±3.41 | 40.05±11.90 | 8.59±2.99 |
| 最大值 | 73.97 | 27.03 | 42.05 | 67.89 | 11.14 |
| 最小值 | 52.10 | 16.60 | 29.32 | 20.45 | 4.16 |
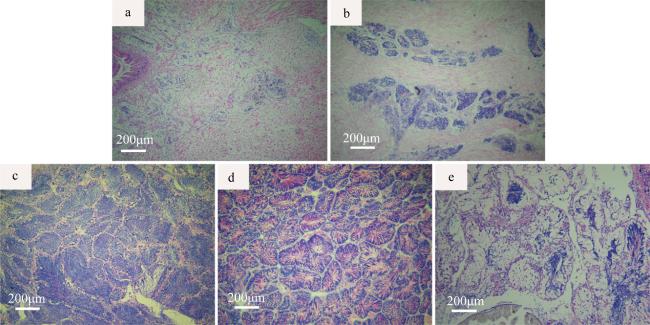
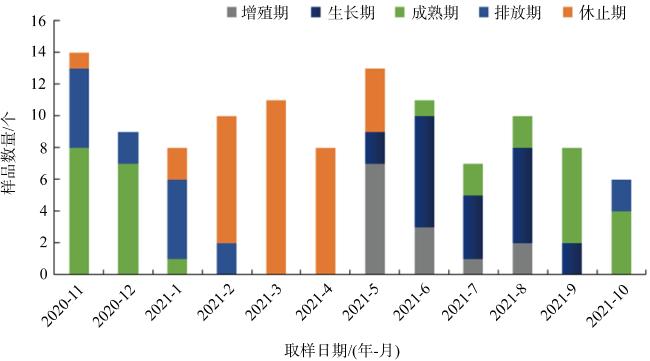
表 2 广西北部湾织锦巴非蛤精巢发育周年分布规律Tab. 2 Annual distribution of spermary development stages of P. textile in the Beibu Gulf, Guangxi |
| 取样日期 | 肥满度 | 精巢发育分期 | 总数/个 | 盐度/‰ | 水温/℃ | ||||
|---|---|---|---|---|---|---|---|---|---|
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | |||||
| 2020-11-29 | 25.20 | 8 | 5 | 1 | 14 | 27.4 | 22.0 | ||
| 2020-12-21 | 21.09 | 7 | 2 | 9 | 27.5 | 17.2 | |||
| 2021-01-08 | 18.33 | 1 | 5 | 2 | 8 | 29.4 | 15.6 | ||
| 2021-02-24 | 16.64 | 2 | 8 | 10 | 30.9 | 17.3 | |||
| 2021-03-18 | 20.26 | 11 | 11 | 30.0 | 21.0 | ||||
| 2021-04-22 | 21.24 | 8 | 8 | 30.0 | 27.5 | ||||
| 2021-05-20 | 27.90 | 7 | 2 | 4 | 13 | 28.0 | 29.5 | ||
| 2021-06-23 | 28.24 | 3 | 7 | 1 | 10 | 28.4 | 30.5 | ||
| 2021-07-15 | 34.89 | 1 | 4 | 2 | 7 | 29.0 | 29.7 | ||
| 2021-08-18 | 34.83 | 2 | 6 | 2 | 10 | 30.0 | 29.8 | ||
| 2021-09-17 | 37.40 | 2 | 6 | 8 | 31.0 | 29.8 | |||
| 2021-10-20 | 33.92 | 4 | 2 | 6 | 29.0 | 25.5 | |||
注: Ⅰ~Ⅴ表格内数字为个体数 |
表3 织锦巴非蛤精细胞各时期发育特点Tab. 3 Development characteristics of the spermatid of P. textile at various stages |
| 发育分期 | 出现时期及位置 | 细胞特点 |
|---|---|---|
| 精原细胞 | 精巢发育各个阶段均可观察到精原细胞, 增殖期较多。由滤泡壁上的上皮细胞增殖而来, 分布在滤泡基膜和生精小管附近。可分为 A型和B型精原细胞。A型细胞一般贴附在滤泡壁或者基膜上, B型分布在滤泡各个部位或紧贴A型精原细胞。 | 精原细胞形状为圆形或椭圆形, 体积在精子发育过程中最大。A型精原细胞胞内物质少、核仁显现, 但不位于细胞中央, 直径在5.04~6.43μm。B型胞内物质趋于紧密、核仁不明显, 直径在4.89~6.98μm。此期精原细胞染色较浅。 |
| 生长初级精母细胞 | 在精巢发育的生长期较多, 成熟期也存在。生长初级精母细胞由精原细胞A型分化而来, 主要分布在生精小管附近, 靠近滤泡壁。 | 形状为椭圆形或者圆形, 直径在3.56~4.78μm, 比精原细胞小, 细胞核不位于细胞的中央。 |
| 生长次级精母细胞 | 生长期和成熟期数量最多, 分布不规律, 在生精小管附近。 | 形状为椭圆形或者圆形, 细胞直径比初级精母细胞小, 直径在2.12~3.87μm, 此期细胞的核膜、核仁不清晰, 细胞核染色质开始浓缩。 |
| 精细胞 | 成熟期、排放期可观察到较多的精细胞, 分布在滤泡中生精小管周围。 | 精细胞分为前期、中期、后期3个时期。前期主要为椭圆形或者圆形, 至中期大部分转变为椭圆形, 后期为成熟精子。此期染色质高度浓缩, 染色程度高。 |
| 成熟精子 | 在成熟期和排放期成熟精子数量最多, 分布在生精小管上, 以头部朝向滤泡壁、尾部聚集成束朝向滤泡腔中心。 | 精细胞经过复杂变化形成具有特殊结构的精子。成熟精子呈子弹头形状, 头部粗、尾部细。染色质高度浓缩, 染色程度深。在滤泡中呈菊花状散布, 聚集成束。 |
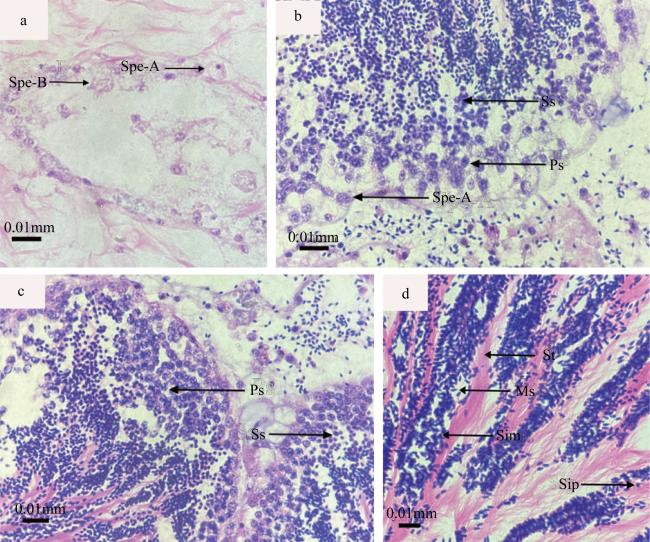
图3 织锦巴非蛤各个时期的雄性生殖细胞a. 精原细胞期, 示A型精原细胞(Spe-A)、B型精原细胞(Spe-B); b. 初级精母细胞期, 示初级精母细胞(Ps)、次级精母细胞(Ss); c. 次级精母细胞期, 示初级精母细胞(Ps)、次级精母细胞(Ss); d. 精细胞期和精子期, 示生精小管(St)、精细胞前期(Sip)、精细胞中期(Sim)、成熟精子(Ms) Fig. 3 The different stages of male reproductive cell of P. textile. (a) Spermatogonium phase, Spe-A: spermatogonia A, Spe-B: spermatogonia B; (b) Primary spermatocyte, Ps: Primary spermatocyte, Ss: Secondary spermatocyte; (c) Secondary spermatocyte phase, Ps: Primary spermatocyte, Ss: Secondary spermatocyte; (d) Sperm cells period and sperm phase, St: Seminiferous tubule, Sip: Spermatid in prophase, Sim; Spermatid in metaphase, Ms: Mature Sperm |
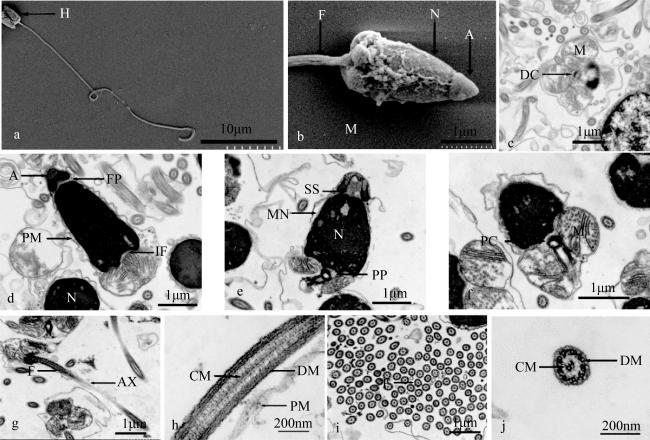
图4 织锦巴非蛤精子的超微结构观察a. 精子, 示头部(H); b. 精子头部和中部, 示顶体(A)、精核(N)、线粒体(M)、鞭毛 (F); c. 精子中部横切, 示远端中心粒(DC)、线粒体(M); d. 精子纵切, 示质膜(PM)、核前窝(FP)、植入窝(IF); e. 精子头部与中部纵切, 示亚顶体腔(SS)、核后窝(PP)、核膜(MN); f. 精子中部纵切, 示近端中心粒(PC)、精核(N)、线粒体(M); g. 尾部鞭毛纵切, 示鞭毛(F)、轴丝(AX); h. 尾部鞭毛纵切, 示中心微管(CM)、双联体微管(DM)、质膜(PM); i. 精子尾部鞭毛横切, 示鞭毛(F); j. 尾部鞭毛横切, 示中心微管(CM)、双联体微管(DM) Fig. 4 Ultrastructural observations on the sperm of P. textile. (a) Sperm, showing head (H); (b) Sperm head and mid-section, showing acrosome (A), nucleus (N), mitochondria (M), flagellum (F); (c) Cross section of mid-section, showing distal centrioles (DC) and mitochondria (M); (d) Sperm longitudinal section, showing plasma membrane (PM), front nuclear pocket (FP), implantation fossa (IF); (e) Longitudinal section of mid-section, showing subacrosomal space (SS), posterior pit (PP), nuclear membranes (NM); (f) Mid-sperm cross section, showing proximal centrioles (PC), nucleus (N); (g) Longitudinal section of tail flagella, showing flagellum (F), axoneme (AX); (h) Longitudinal section of tail flagella, showing central microtubule (CM), doublet microtubules (DM), plasm membrane (PM); (i) Cross section of tail flagella, showing flagellum (F); (j) Cross section of tail flagella, showing microtubule (CM), doublet microtubules (DM) |
表4 不同海产双壳贝类精子超微结构比较Tab. 4 Ultrastructural comparison of the sperm of different marine bivalve shellfish |
| 物种 | 精子长度/μm | 顶体形状 | 精核长宽比值 | 线粒体数目 | 尾部结构 | 参考文献 | |
|---|---|---|---|---|---|---|---|
| 织锦巴非蛤(Paphia textile) 近江蛏(Sinonovacula rivularis) | 46.04 57.00~58.00 | 圆锥形 保龄球形 | 1.47 0.36 | 4 4 | “9+2”双联体微管结构 “9+2”双联体微管结构 | 本文 黄瑞等(2011) | |
| 钝缀锦蛤(Tapes conspersus) | 59.27 | 倒“V”形 | 1.27 | 5 | “9+2”双联体微管结构 | 连昌朋等(2022) | |
| 靓巴非蛤(Paphia schnelliana) 施氏獭蛤(Lutraria sieboldii) | 50.16 73.21 | 圆锥形 半球形 | 1.42 1.14 | 4 5 | — — | Chen等(2021) Chen等(2021) | |
| 魁蚶(Scapharca broughtoni) | — | 圆锥形 | 0.89 | 5 | “9+2”双联体微管结构 | 叶婧等(2012) | |
| 斧文蛤(Meretrix lamarkii) | 45.20~47.70 | 圆锥形 | 1.41 | 5 | “9+2”双联体微管结构 | 董迎辉等(2012) | |
| 波纹巴非蛤(Paphia undulata) 红树蚬(Polymesoda erosa) | — — | 圆锥形 倒“V”形 | 1.43 4.00 | 4 4 | “9+2”双联体微管结构 “9+2”双联体微管结构 | 赵志江等(1992) 郭云鹏等(2020) | |
| 长牡蛎(Crassostrea gigas) | 57.77 | 圆锥形 | 1.12 | 4 | “9+2”双联体微管结构 | Dong等(2005) | |
| 虾夷扇贝(Patinopecten yessoensis) | 50.00 | 倒“V”形 | 3.00 | 4~5 | “9+2”双联体微管结构 | 韩厚伟等(2008) | |
注: —表示暂无数据 |
| [1] |
邓道贵, 谈奇坤, 2000. 褶纹冠蚌精子发生的研究[J]. 水生生物学报, 24(1): 63-66.
|
| [2] |
邓正华, 翟子钦, 魏海军, 等, 2022. 织锦巴非蛤幼虫对不同种类单胞藻的摄食和消化效果[J]. 南方农业学报, 53(5): 1448-1556.
|
| [3] |
董迎辉, 林志华, 姚韩韩, 等, 2012. 斧文蛤精子超微结构与受精过程的细胞学变化[J]. 水产学报, 35(3): 356-364.
|
| [4] |
郭延平, 谈奇坤, 陈士超, 2002. 三角帆蚌精子的形态及超微结构[J]. 动物学杂志, 37(2): 10-13.
|
| [5] |
郭云鹏, 王公嗣, 黄勃, 2020. 红树蚬精巢发育的组织学研究和精子超微结构的观察[J]. 福建农林大学学报(自然科学版), 49(4): 519-523.
|
| [6] |
韩厚伟, 高悦勉, 刘春凤, 等, 2008. 虾夷扇贝精子的超微结构[J]. 动物学杂志, 43(1): 75-81.
|
| [7] |
何义朝, 张福绥, 王萍, 等, 1999. 墨西哥湾扇贝稚贝对盐度的耐受力[J]. 海洋学报, 21(4): 87-91.
|
| [8] |
黄洪龙, 骆轩, 柯才焕, 2021. 腹足类精子超微结构研究进展[J]. 渔业研究, 43(1): 103-110.
|
| [9] |
黄瑞, 黄标武, 李林春, 等, 2011. 近江蛏精子超微形态结构观察及与缢蛏精子的比较[J]. 水产学报, 35(1): 58-65.
|
| [10] |
姜明, 汝少国, 陶迺蓉, 等, 2003. 入卵过程中太平洋牡蛎精子超微结构的变化[J]. 青岛海洋大学学报, 33(4): 551-556.
|
| [11] |
赖胜琪, 尹聪, 邱炬维, 等, 2022. 北部湾沿海织锦巴非蛤不同地理群体形态差异研究[J]. 广东农业科学, 49(7): 105-112.
|
| [12] |
栗志民, 刘志刚, 韩伟贤, 等, 2011. 织锦巴非蛤稚贝盐度适应性研究[J]. 海洋科学, 35(10): 96-102.
|
| [13] |
连昌朋, 王超奇, 杨凌, 等, 2022. 广西北部湾钝缀锦蛤精巢发育、精子发生及超微结构研究[J]. 海洋科学, 46(6): 80-89.
|
| [14] |
刘海娟, 陈瑞芳, 曾梦清, 等, 2022. 织锦巴非蛤人工养殖技术研究[J]. 科学养鱼, 38(11): 61-62.
|
| [15] |
鹿瑶, 刘辉, 聂鸿涛, 等, 2015. 辽宁沿海薄片镜蛤的繁殖周期研究[J]. 大连海洋大学学报, 30(6): 647-652.
|
| [16] |
宁军号, 常亚青, 宋坚, 等, 2015. 偏顶蛤的性腺发育和生殖周期[J]. 中国水产科学, 22(3): 469-477.
|
| [17] |
吴加莹, 戴明姝, 刘志刚, 等, 2023. 温度对织锦巴非蛤稚贝生存和生长的影响[J]. 南方水产科学, 19(2): 1-8.
|
| [18] |
徐炳杰, 刘一鸣, 连昌朋, 等, 2024. 广西北部湾海域织锦巴非蛤卵巢发育、卵子及卵黄发生的研究[J]. 热带海洋学报, https://kns.cnki.net/kcms2/detail/44.1500.P.20230726.0854.002.html.
|
| [19] |
徐凤山, 张素萍, 2008. 中国海产双壳类图志[M]. 北京: 科学出版社:244- 245.
|
| [20] |
叶婧, 姜建湖, 2012. 魁蚶精子发生的超微结构[J]. 上海海洋大学学报, 21(2): 199-203.
|
| [21] |
尤仲杰, 陆彤霞, 马斌, 等, 2003. 几种环境因子对墨西哥湾扇贝幼虫和稚贝生长与存活的影响[J]. 热带海洋学报, 22(3): 22-29.
|
| [22] |
余红卫, 2012. 彩虹明樱蛤精子发生的超微结构[J]. 电子显微学报, 31(1): 65-69.
|
| [23] |
翟子钦, 喻达辉, 白丽蓉, 2022. 织锦巴非蛤形态性状对体质量的影响[J]. 广东农业科学, 49(7): 113-119.
|
| [24] |
张福绥, 马江虎, 何义朝, 等, 1991. 胶州湾海湾扇贝肥满度的研究[J]. 海洋与湖沼, 22(2): 97-103.
|
| [25] |
赵志江, 李复雪, 柯才焕, 1991. 波纹巴非蛤的性腺发育和生殖周期[J]. 水产学报, 15(1): 1-8.
|
| [26] |
赵志江, 李复雪, 1992. 波纹巴非蛤Paphia undulata精子发生的超微结构[J]. 台湾海峡, 11(3): 238-243.
|
| [27] |
郑学斌, 张清科, 乐韵, 等, 2018. 香鱼(Plecoglossus altivelis)精子的超微结构及其与鲤形目及鲑形目其他鱼类精子结构的比较研究[J]. 海洋与湖沼, 49(4): 866-872.
|
| [28] |
周丽青, 杨爱国, 刘志鸿, 等, 2011. 扇贝精子及卵子的受精生物学特性[J]. 渔业科学进展, 32(1): 75-81.
|
| [29] |
周小龙, 董迎辉, 边平江, 等, 2012. 帘文蛤精子超微结构及与其他双壳贝类的比较[J]. 台湾海峡, 31(4): 495-500.
|
| [30] |
竺俊全, 杨万喜, 2002. 双壳类软体动物精子发生及其在系统演化研究中的应用前景[J]. 海洋湖沼通报, 4(5): 25-31.
|
| [31] |
朱星海, 孙红振, 杨祖晶, 等, 2019. 风信标扇贝的性腺发育与繁殖周期规律研究[J]. 中国海洋大学学报(自然科学版), 49(2): 52-58.
|
| [32] |
庄启谦, 2001. 中国动物志软体动物门双壳纲帘蛤科[M]. 北京:科学出版社:53- 54.
|
| [33] |
邹杰, 彭慧婧, 杨家林, 2019. 织锦巴非蛤人工种苗培育及浅海养殖试验[J]. 科学养鱼, 35(10): 57-58.
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}