
潮滩冲淤扰动下外来与乡土红树植物幼苗稳定性差异
|
潘炜杰(1992—), 男, 广东省佛山市人, 硕士研究生, 从事滨海湿地保护研究。email: |
Editor: 林强
收稿日期: 2020-11-12
修回日期: 2020-02-27
网络出版日期: 2021-03-10
基金资助
广东省重点实验室专项(2019B121203011)
南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0403)
版权
Performance of exotic and native mangrove seedlings under accretion/erosion disturbances on tidal flats
Editor: LIN Qiang
Received date: 2020-11-12
Revised date: 2020-02-27
Online published: 2021-03-10
Supported by
Guangdong Provincial Key Laboratory Project(2019B121203011)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0403)
Copyright
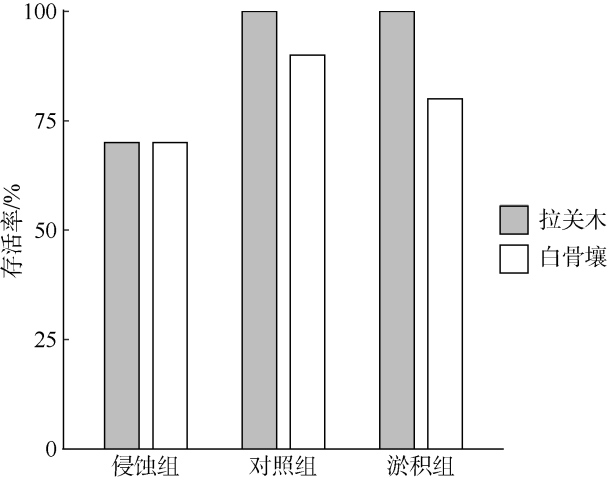
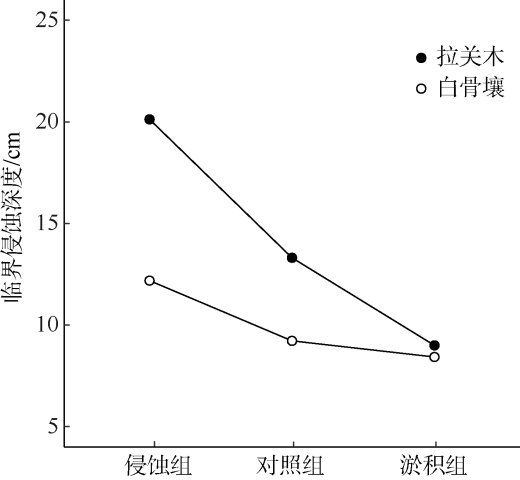
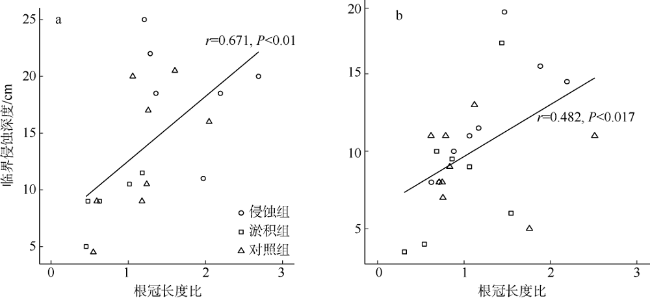
引种外来树种是我国各地在红树林修复实践中常用的措施。由于更容易在困难立林地存活, 外来树种被认为更具竞争优势。为弄清外来与乡土红树植物物种在困难立林地存活能力的差异及其原因, 本研究选取典型外来引种先锋树种拉关木(Laguncularia racemosa)与乡土先锋植物白骨壤(Avicennia marina)为研究对象, 通过控制实验探究潮滩冲淤扰动条件下幼苗稳定性差异及其形成机制。结果表明, 幼苗抗侵蚀能力与幼苗根冠比显著正相关, 泥沙淤积抑制幼苗根系生长, 而底床侵蚀则促进根系伸长。与白骨壤相比, 拉关木幼苗的主根更长, 根冠长度比更大, 更能抵抗底泥的侵蚀扰动, 且通过根系伸长以增强幼苗稳定性的能力也更强。拉关木幼苗比白骨壤更能适应潮滩冲淤扰动, 其竞争优势从幼苗阶段就已经建立。本研究结果可为红树林修复实践中树种和宜林地的选择提供参考, 亦可为评估外来红树物种入侵风险评价提供依据。
潘炜杰 , 祝振昌 , 蔡宴朋 , 杨志峰 . 潮滩冲淤扰动下外来与乡土红树植物幼苗稳定性差异[J]. 热带海洋学报, 2021 , 40(6) : 120 -127 . DOI: 10.11978/2020133
Exotic species are only used in some places in China for mangrove restoration, as they often perform better than native species on these desired restoration sites often with highly stressful habitats. Here, we conducted a mesocosm experiment to detect the differences of survivability between exotic and native mangrove seedlings under accretion/erosion disturbances on tidal flats, using exotic species Laguncularia racemosa and native pioneer species Avicennia marina as examples. Our results demonstrated that for both species the erosion resistance of seedlings correlated positively with root-to-shoot length ratio. Sediment accretion disturbance inhibited the root development of seedlings so that the root-to-shoot length ratio increased, while sediment erosion promoted root elongation with the root-to-shoot length ratio decreased. Compared with the native species A. marina, exotic species L. racemosa seedlings have greater root-to-shoot ratio, and are thus more resistant to erosion with better adaptability to sediment dynamics via root elongation. Our study indicates that the exotic species L. racemosa are more adaptable to erosion disturbances on tidal flats than the native species A. marina. These findings shed light on the selection of tree species and suitable planting sites for mangrove restoration, and might provide helpful insights into assessing the risk of species invasion in mangrove ecosystems.
Key words: Accretion/erosion; Laguncularia racemosa; Avicennia marina; seedling; stability
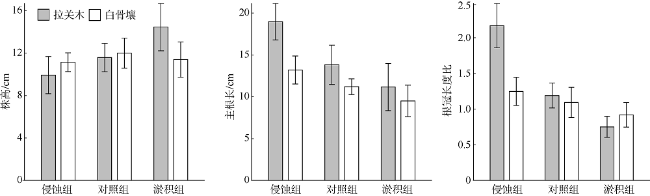
图3 冲淤作用对幼苗株高(a)和主根长(b)的影响及各组根冠长度比(c)Fig. 3 Effects of simulated sediment dynamics on seedlings grown (a) and main root length (b) and the root-to-shoot length ratio (c) of each group |
表1 物种类型和冲淤处理对幼苗株高、主根长与临界侵蚀深度影响的双因素方差分析Tab. 1 Two-way ANOVAs of the effects of accretion/erosion treatment and species type on the shoot height, length of main root and critical erosion depth, respectively |
| 响应变量 | 差异源 | d.f. | F value | Pr(>F) |
|---|---|---|---|---|
| 株高 | 处理 | 2 | 0.951 | 0.395 |
| 物种 | 1 | 0.006 | 0.939 | |
| 处理: 物种 | 2 | 0.967 | 0.389 | |
| 主根长 | 处理 | 2 | 4.03 | 0.0257 |
| 物种 | 1 | 5.642 | 0.0225 | |
| 处理: 物种 | 2 | 0.598 | 0.5546 | |
| 根冠长度比 | 处理 | 2 | 7.093 | <0.001 |
| 物种 | 1 | 4.079 | 0.05 | |
| 处理: 物种 | 2 | 3.226 | 0.051 | |
| 临界侵蚀深度 | 处理 | 2 | 15.297 | <0.001 |
| 物种 | 1 | 10.819 | <0.001 | |
| 处理: 物种 | 2 | 2.642 | 0.083 |
| [1] |
陈坚, 李妮亚, 刘强, 等, 2013. NaCl处理下两种引进红树的光合及抗氧化防御能力[J]. 植物生态学报, 37(5):443-453.
|
| [2] |
陈鹭真, 王文卿, 张宜辉, 等, 2010. 2008年南方低温对我国红树植物的破坏作用[J]. 植物生态学报, 34(2):186-194.
|
| [3] |
邓必玉, 吴玲巧, 秦旭东, 等, 2020. 广西红树林主要外来植物现状及防控对策研究[J]. 林业调查规划, 45(4):54-60.
|
| [4] |
范航清, 王文卿, 2017. 中国红树林保育的若干重要问题[J]. 厦门大学学报(自然科学版), 56(3):323-330.
|
| [5] |
廖宝文, 李玫, 陈玉军, 等, 2010. 中国红树林恢复与重建技术[M]. 北京: 科学出版社.
|
| [6] |
李天杰, 赵烨, 张科利, 等, 2004. 土壤地理学[M]. 3版. 北京: 高等教育出版社.
|
| [7] |
林天维, 柴清志, 孙子钧, 等, 2020. 我国红树林的面积变化及其治理[J]. 海洋开发与管理, 37(2):48-52.
|
| [8] |
卢昌义, 廖宝文, 2019. 对外来红树植物无瓣海桑和拉关木生态作用的思考[J]. 湿地科学, 17(6):682-688.
|
| [9] |
谭芳林, 卢昌义, 林捷, 等, 2018. 福建省外来红树植物引种及扩散状况调研报告[J]. 福建林业, (4):28-33 (in Chinese).
|
| [10] |
王炳宇, 杨珊, 刘强, 等, 2020. 外来红树植物无瓣海桑和拉关木在海南东寨港的人工种植与自然扩散[J]. 生态学杂志, 39(6):1778-1786.
|
| [11] |
王秀丽, 卢昌义, 周亮, 等, 2017a. 外来红树植物拉关木对木榄的化感作用[J]. 厦门大学学报(自然科学版), 56(3):339-345.
|
| [12] |
王秀丽, 周亮, 卢昌义, 2017b. 从木榄叶片超微结构所受的影响评估外来种拉关木的化感作用[J]. 生态科学, 36(5):177-185.
|
| [13] |
王友绍, 红树林分子生态学[M]. 2019. 北京: 科学出版社.
|
| [14] |
向敏, 刘强, 李妮亚, 等, 2016. 引进红树拉关木和两种乡土红树离子平衡及光合作用的比较研究[J]. 广西植物, 36(4):387-396.
|
| [15] |
杨珊, 刘强, 王炳宇, 等, 2020. 外来红树植物拉关木对乡土种桐花树和正红树的化感作用研究[J]. 广西植物, 40(3):356-366.
|
| [16] |
张苇, 廖宝文, 刘滨尔, 2013. 水东湾困难立地滩涂红树林生长表现分析[J]. 防护林科技, (11):1-3.
|
| [17] |
钟才荣, 李诗川, 杨宇晨, 等, 2011. 红树植物拉关木的引种效果调查研究[J]. 福建林业科技, 38(3):96-99.
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}