
短指软珊瑚(Sinularia acuta)热休克蛋白HSP70家族特征及进化分析
|
李子若(2001—), 女, 海南乐东县人, 大学本科, 从事海洋生物遗传多样性研究。email: |
Copy editor: 孙翠慈
收稿日期: 2023-09-17
修回日期: 2023-10-07
网络出版日期: 2023-11-29
基金资助
海南省重点研发项目(ZDYF2021SHFZ104)
省级大学生创新创业训练计划项目(S202111100016)
中华环境保护基金会美团外卖青山公益专项基金(CEPFQS202169-27)
Predicted family characteristics and evolution of the heat shock protein HSP70 in Sinularia acuta
Copy editor: SUN Cuici
Received date: 2023-09-17
Revised date: 2023-10-07
Online published: 2023-11-29
Supported by
Key Research and Development Project of Hainan Province(ZDYF2021SHFZ104)
Provincial Student Innovation and Entrepreneurship Training Program(S202111100016)
Special Fund for Green Mountain Public Welfare of Meituan Takeout of China Environmental Protection Foundation(CEPFQS202169-27)
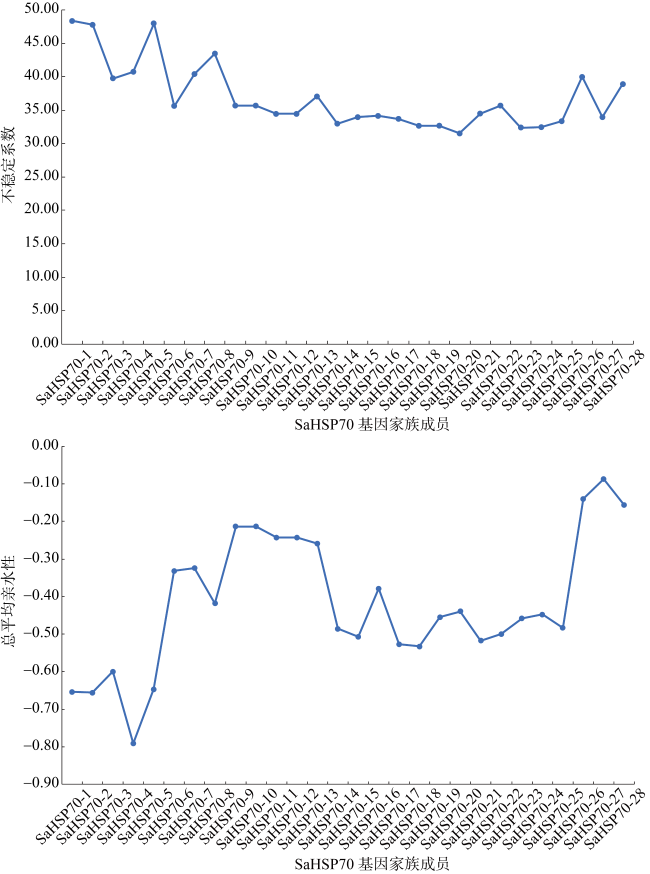
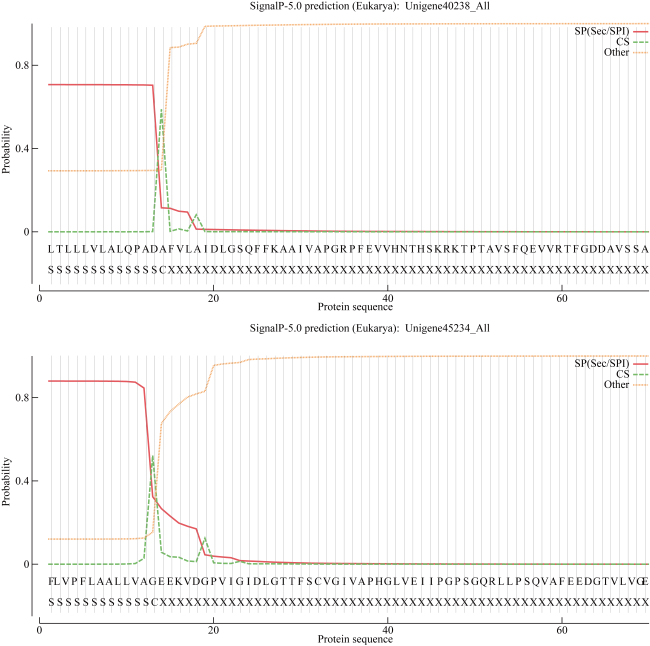
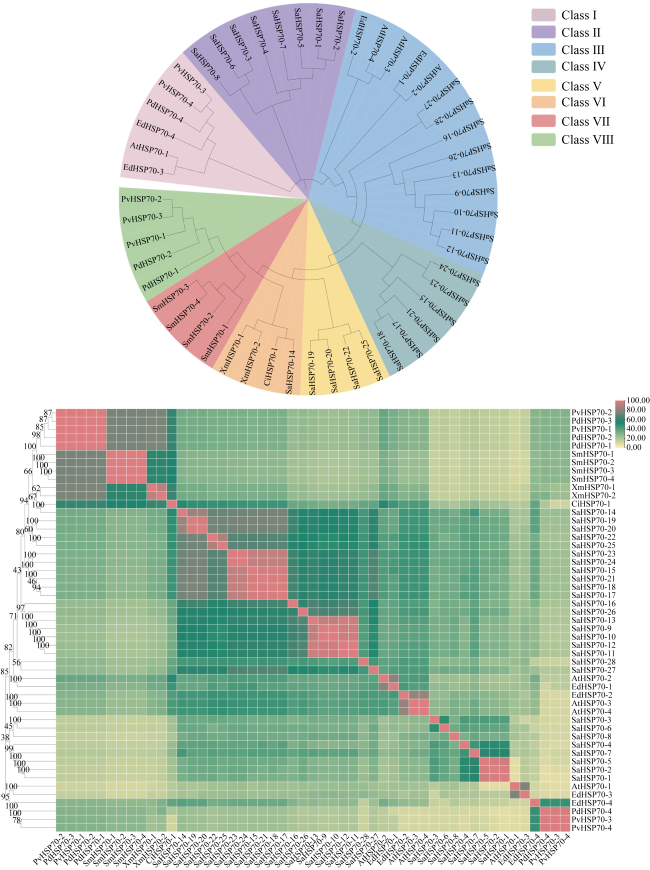
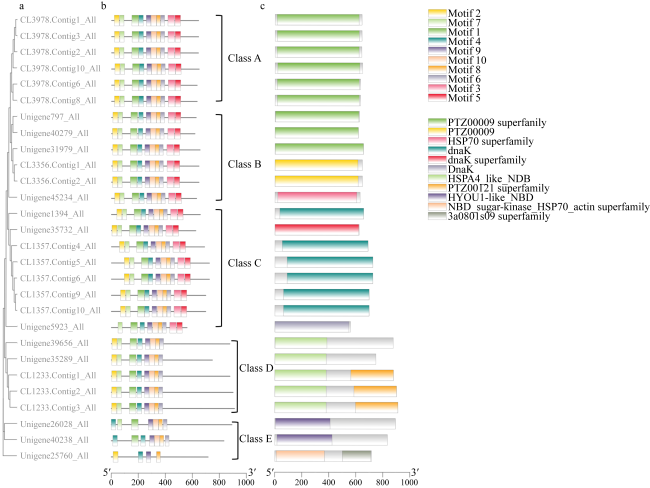
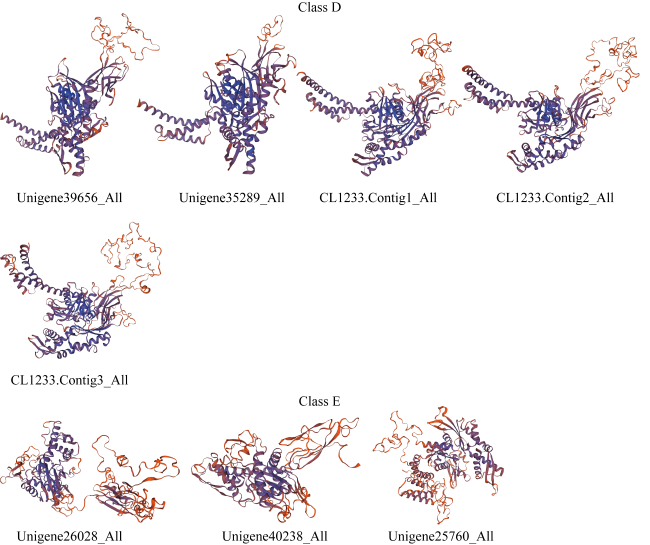
热休克蛋白70(heat shock protein 70, HSP70)在生物细胞或组织免受热或氧化应激等方面起着关键作用, 是已知高度保守的蛋白质之一。由于全球环境持续升温, 珊瑚大面积白化、死亡, 珊瑚如何应对持续升温的抗逆机制是科学研究热点。本研究从高温胁迫短指软珊瑚测序蛋白序列数据库分析鉴定出了28个HSP70蛋白家族成员, 均为酸性亲水蛋白, 大部分蛋白质结构较为稳定。亚细胞定位表明HSP70蛋白主要分布在珊瑚细胞核、细胞质中, 在线粒体、内质网上也有少量分布。信号肽预测表明, 28个HSP70蛋白成员中26个没有信号肽, 大部分不属于分泌蛋白, 不存在跨膜结构。系统进化树结果表明短指软珊瑚HSP70蛋白家族成员聚成5大类。短指软珊瑚HSP70蛋白家族结构和保守基序分析中预测到了10条保守基序motif分为5个亚族。短指软珊瑚HSP70蛋白家族二级结构主要以α-螺旋和无规则卷曲为主, α-螺旋的含量占比大。28个HSP70家族蛋白中有25个预测到了N-糖基化位点, 且位点个数在1~9范围内。28个HSP70家族蛋白均预测到磷酸化位点和O-糖基化位点, 总个数分别在41~96和1~23范围内。本研究HSP70家族蛋白结果为今后珊瑚在应对全球升温胁迫中的适应机制等方面研究奠定了基础。
李子若 , 罗晏杰 , 曹政 , CHIN Yaoxian , 王沛政 . 短指软珊瑚(Sinularia acuta)热休克蛋白HSP70家族特征及进化分析[J]. 热带海洋学报, 2024 , 43(4) : 123 -136 . DOI: 10.11978/2023134
Heat shock protein 70 (HSP70) is a highly conserved protein that plays a key role in protecting biological cells or tissues from heat or oxidative stress. Due to the continuous warming of the global environment and the widespread bleaching and death of corals, how corals respond to the stress resistance mechanism of continuous warming is a scientific research hotspot. However, current research on the characteristics of the HSP70 family in corals are relatively rare. In this study, 28 members of the HSP70 family were identified by analysis of the Sinularia acuta protein sequences under high temperature stress, all of which were acidic hydrophilic proteins with relatively stable structure. Subcellular localization showed that HSP70 was mainly found in coral nuclei and cytoplasm, with lesser amount in mitochondria and endoplasmic reticulum. Signal peptide predictions showed that 26 of the 28 HSP70 members did not have signal peptides, most of them do not secrete proteins and did not have a transmembrane structure. The phylogenetic tree showed that the members of the S. acuta HSP70 family can be clustered into 5 classes. Analysis of the structure and conserved regions of the hsp70 gene family of S. acuta predicted 10 motifs which could be divided into 5 subfamilies. The main secondary structure of the HSP70 protein in S. acuta was α-helix followed by random coiling. N-glycosylation sites were predicted in 25 of the 28 HSP70 family proteins, with the number of sites ranging from 1 to 9. All 28 HSP70 family proteins were predicted to have phosphorylation and O-glycosylation sites, with a total number in the range of 41~96 and 1~23, respectively. The results of this study on the HSP70 family lay the foundation for future research on the adaptive mechanisms of corals in response to global warming stress.
Key words: Sinularia acuta; heat shock proteins; HSP70 family; evolution
表1 分析工具网址及功能Tab. 1 Website and use of analysis tools |
| 分析工具 | 网址 | 功能 |
|---|---|---|
| NCBI CD-search | https://www.ncbi.nlm.nih.gov/cdd/ | 保守结构域预测 |
| ExPASy-ProtParam tool | https://web.expasy.org/protparam/ | 蛋白质的氨基酸数量、分子量、等电点、稳定性 |
| Singa IP 5.0 | https://services.healthtech.dtu.dk/service.php?SignalP-5.0 | 信号肽预测 |
| ExPASy-ProtScale tool | https://www.expasy.org/resources/protscale | 蛋白质的亲水性 |
| TMHMM 2.0 | https://services.healthtech.dtu.dk/service.php?TMHMM-2.0 | 跨膜区域预测 |
| WoLF PSORT | https://wolfpsort.hgc.jp/ | 亚细胞定位预测 |
| MEME | https://meme-suite.org/meme/tools/meme | 保守结构域分析 |
| SWISS-MODEL | https://swissmodel.expasy.org | 蛋白质三维结构 |
| SOPMA | http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html | 蛋白质的二级结构 |
| NetPhos 3.1 | https://services.healthtech.dtu.dk/service.php?NetPhos-3.1 | 磷酸化位点预测 |
| NetNGlyc 1.0 | https://services.healthtech.dtu.dk/service.php?NetNGlyc-1.0 | N-糖基化位点预测 |
| NetOGlyc 4.0 | https://services.healthtech.dtu.dk/service.php?NetOGlyc-4.0 | O-糖基化位点预测 |
表2 短指软珊瑚HSP70家族信息Tab. 2 Information on HSP70 family in S. acuta |
| 基因编号 | 蛋白质ID | 氨基酸数量 | 分子量 | 等电点 | 亚细胞定位 |
|---|---|---|---|---|---|
| aHSP70-1 | CL1233.Contig3_All | 912 | 101494.21 | 4.82 | 细胞核 |
| SaHSP70-2 | CL1233.Contig2_All | 901 | 100456.04 | 4.80 | 细胞核 |
| SaHSP70-3 | Unigene26028_All | 893 | 100307.02 | 5.87 | 细胞核 |
| SaHSP70-4 | Unigene39656_All | 877 | 99089.04 | 5.12 | 细胞质膜和细胞核膜 |
| SaHSP70-5 | CL1233.Contig1_All | 878 | 98371.94 | 4.87 | 细胞核 |
| SaHSP70-6 | Unigene40238_All | 834 | 92628.05 | 5.22 | 内质网 |
| SaHSP70-7 | Unigene35289_All | 748 | 82074.38 | 5.31 | 细胞核、细胞质 |
| SaHSP70-8 | Unigene25760_All | 717 | 79097.83 | 5.36 | 细胞质膜和细胞核膜 |
| SaHSP70-9 | CL1357.Contig5_All | 725 | 76099.19 | 5.03 | 线粒体 |
| SaHSP70-10 | CL1357.Contig6_All | 725 | 76099.19 | 5.03 | 线粒体 |
| SaHSP70-11 | CL1357.Contig10_All | 698 | 73525.14 | 4.92 | 细胞质 |
| SaHSP70-12 | CL1357.Contig9_All | 698 | 73525.14 | 4.92 | 细胞质 |
| SaHSP70-13 | CL1357.Contig4_All | 689 | 72820.29 | 4.91 | 细胞质 |
| SaHSP70-14 | Unigene31979_All | 655 | 71933.13 | 5.20 | 细胞质 |
| SaHSP70-15 | CL3978.Contig10_All | 648 | 71104.23 | 5.07 | 细胞质 |
| SaHSP70-16 | Unigene1394_All | 657 | 70957.30 | 5.18 | 线粒体 |
| SaHSP70-17 | CL3978.Contig3_All | 643 | 70673.78 | 5.12 | 细胞质 |
| SaHSP70-18 | CL3978.Contig1_All | 643 | 70638.72 | 5.12 | 细胞质 |
| SaHSP70-19 | CL3356.Contig1_All | 647 | 70578.80 | 5.08 | 细胞质 |
| SaHSP70-20 | CL3356.Contig2_All | 647 | 70562.86 | 5.13 | 细胞质 |
| SaHSP70-21 | CL3978.Contig2_All | 642 | 70453.51 | 5.12 | 细胞质 |
| SaHSP70-22 | Unigene797_All | 626 | 70077.35 | 5.37 | 细胞质 |
| SaHSP70-23 | CL3978.Contig6_All | 633 | 69758.80 | 5.03 | 细胞质 |
| SaHSP70-24 | CL3978.Contig8_All | 633 | 69666.51 | 4.96 | 细胞质 |
| SaHSP70-25 | Unigene40279_All | 617 | 68987.59 | 5.98 | 细胞质 |
| SaHSP70-26 | Unigene35732_All | 624 | 67397.85 | 4.98 | 细胞核、细胞质 |
| SaHSP70-27 | Unigene45234_All | 630 | 67279.60 | 5.22 | 细胞质基质外 |
| SaHSP70-28 | Unigene5923_All | 560 | 61929.42 | 5.92 | 细胞质 |
表3 暗色海葵等7个参考物种的信息来源Tab. 3 Information on seven species of Actinia tenebrosa etc |
| 物种中文名 | 物种拉丁文名 | 基因登录号 | 编号 |
|---|---|---|---|
| 暗色海葵 | Actinia tenebrosa | XP_031562268.1 | AtHSP70-1 |
| Actinia tenebrosa | XP_031555558.1 | AtHSP70-2 | |
| Actinia tenebrosa | XP_031552530.1 | AtHSP70-3 | |
| Actinia tenebrosa | XP_031552529.1 | AtHSP70-4 | |
| 海葵 | Exaiptasia diaphana | KXJ22228.1 | EdHSP70-1 |
| Exaiptasia diaphana | KXJ18030.1 | EdHSP70-2 | |
| Exaiptasia diaphana | KXJ15022.1 | EdHSP70-3 | |
| Exaiptasia diaphana | KXJ09058.1 | EdHSP70-4 | |
| 鹿角杯形珊瑚 | Pocillopora damicornis | AFW20530.1 | PdHSP70-1 |
| Pocillopora damicornis | AFW20529.1 | PdHSP70-2 | |
| Pocillopora damicornis | AFW20516.1 | PdHSP70-3 | |
| Pocillopora damicornis | AFW20572.1 | PdHSP70-4 | |
| 疣状杯形珊瑚 | Pocillopora verrucosa | AFW20524.1 | PvHSP70-1 |
| Pocillopora verrucosa | AFW20525.1 | PvHSP70-2 | |
| Pocillopora verrucosa | AFW20582.1 | PvHSP70-3 | |
| Pocillopora verrucosa | AFW20581.1 | PvHSP70-4 | |
| 柱形珊瑚 | Stylophora mamillata | CZF87364.1 | SmHSP70-1 |
| Stylophora mamillata | CZF87363.1 | SmHSP70-2 | |
| Stylophora mamillata | CZF87365.1 | SmHSP70-3 | |
| Stylophora mamillata | CZF87360.1 | SmHSP70-4 | |
| 桶状海绵 | Xestospongia muta | ABY54994.1 | XmHSP70-1 |
| Xestospongia muta | ABY54993.1 | XmHSP70-2 | |
| 软珊瑚 | Capnella imbricata | AJA72720.1 | CiHSP70-1 |
表4 软珊瑚HSP70蛋白的二级结构预测Tab. 4 Secondary structure prediction of HSP70 |
| 基因ID | 基因命名 | α-螺旋/% | β-折叠/% | 延伸链/% | 无规则卷曲/% |
|---|---|---|---|---|---|
| SaHSP70-1 | CL1233.Contig3_All | 45.72 | 4.06 | 13.71 | 36.51 |
| SaHSP70-2 | CL1233.Contig2_All | 45.28 | 4.00 | 13.54 | 37.18 |
| SaHSP70-3 | Unigene26028_All | 49.61 | 3.70 | 12.43 | 34.27 |
| SaHSP70-4 | Unigene39656_All | 44.47 | 3.08 | 13.34 | 39.11 |
| SaHSP70-5 | CL1233.Contig1_All | 48.86 | 3.30 | 12.87 | 34.97 |
| SaHSP70-6 | Unigene40238_All | 48.92 | 3.48 | 14.03 | 33.57 |
| SaHSP70-7 | Unigene35289_All | 51.74 | 3.21 | 14.44 | 30.61 |
| SaHSP70-8 | Unigene25760_All | 64.99 | 1.67 | 9.07 | 24.27 |
| SaHSP70-9 | CL1357.Contig5_All | 42.62 | 6.90 | 19.72 | 30.76 |
| SaHSP70-10 | CL1357.Contig6_All | 42.62 | 6.90 | 19.72 | 30.76 |
| SaHSP70-11 | CL1357.Contig10_All | 42.41 | 6.45 | 19.91 | 31.23 |
| SaHSP70-12 | CL1357.Contig9_All | 42.41 | 6.45 | 19.91 | 31.23 |
| SaHSP70-13 | CL1357.Contig4_All | 42.53 | 6.79 | 20.17 | 30.33 |
| SaHSP70-14 | Unigene31979_All | 41.68 | 7.33 | 17.86 | 33.13 |
| SaHSP70-15 | CL3978.Contig10_All | 43.67 | 7.41 | 18.98 | 29.94 |
| SaHSP70-16 | Unigene1394_All | 40.94 | 7.46 | 21.46 | 30.14 |
| SaHSP70-17 | CL3978.Contig3_All | 42.30 | 7.78 | 20.06 | 29.86 |
| SaHSP70-18 | CL3978.Contig1_All | 44.01 | 6.84 | 19.44 | 29.70 |
| SaHSP70-19 | CL3356.Contig1_All | 42.04 | 8.50 | 17.93 | 31.53 |
| SaHSP70-20 | CL3356.Contig2_All | 42.04 | 8.66 | 18.08 | 31.22 |
| SaHSP70-21 | CL3978.Contig2_All | 42.83 | 7.48 | 20.09 | 29.60 |
| SaHSP70-22 | Unigene797_All | 43.13 | 7.03 | 18.85 | 30.99 |
| SaHSP70-23 | CL3978.Contig6_All | 42.02 | 6.79 | 19.27 | 31.91 |
| SaHSP70-24 | CL3978.Contig8_All | 42.50 | 7.74 | 19.59 | 30.17 |
| SaHSP70-25 | Unigene40279_All | 43.44 | 6.32 | 19.12 | 31.12 |
| SaHSP70-26 | Unigene35732_All | 44.23 | 6.89 | 20.35 | 28.53 |
| SaHSP70-27 | Unigene45234_All | 42.70 | 7.62 | 19.05 | 30.63 |
| SaHSP70-28 | Unigene5923_All | 35.36 | 8.04 | 22.68 | 33.93 |
表5 HSP70磷酸化位点预测Tab. 5 Phosphorylation, N-Glycosylation and O-Glycosylation sites prediction of HSP70 protein |
| 序号 | 基因号 | 丝氨酸 | 苏氨酸 | 酪氨酸 | N-糖基化位点 | O-糖基化位点 |
|---|---|---|---|---|---|---|
| SaHSP70-1 | CL1233.Contig3_All | 38 | 31 | 5 | 3 | 12 |
| SaHSP70-2 | CL1233.Contig2_All | 37 | 29 | 5 | 2 | 10 |
| SaHSP70-3 | Unigene26028_All | 56 | 33 | 7 | 8 | 21 |
| SaHSP70-4 | Unigene39656_All | 42 | 27 | 8 | 3 | 23 |
| SaHSP70-5 | CL1233.Contig1_All | 37 | 29 | 5 | 2 | 7 |
| SaHSP70-6 | Unigene40238_All | 42 | 23 | 9 | 9 | 7 |
| SaHSP70-7 | Unigene35289_All | 39 | 23 | 5 | 2 | 6 |
| SaHSP70-8 | Unigene25760_All | 37 | 17 | 6 | 0 | 3 |
| SaHSP70-9 | CL1357.Contig5_All | 32 | 27 | 3 | 4 | 20 |
| SaHSP70-10 | CL1357.Contig6_All | 31 | 27 | 3 | 4 | 20 |
| SaHSP70-11 | CL1357.Contig10_All | 30 | 26 | 3 | 4 | 13 |
| SaHSP70-12 | CL1357.Contig9_All | 28 | 26 | 2 | 4 | 13 |
| SaHSP70-13 | CL1357.Contig4_All | 30 | 23 | 3 | 4 | 14 |
| SaHSP70-14 | Unigene31979_All | 25 | 28 | 6 | 6 | 7 |
| SaHSP70-15 | CL3978.Contig10_All | 24 | 20 | 9 | 1 | 1 |
| SaHSP70-16 | Unigene1394_All | 33 | 24 | 3 | 4 | 11 |
| SaHSP70-17 | CL3978.Contig3_All | 23 | 21 | 7 | 1 | 3 |
| SaHSP70-18 | CL3978.Contig1_All | 23 | 21 | 9 | 1 | 1 |
| SaHSP70-19 | CL3356.Contig1_All | 24 | 27 | 7 | 5 | 2 |
| SaHSP70-20 | CL3356.Contig2_All | 21 | 26 | 6 | 4 | 3 |
| SaHSP70-21 | CL3978.Contig2_All | 23 | 22 | 7 | 1 | 2 |
| SaHSP70-22 | Unigene797_All | 24 | 17 | 6 | 4 | 3 |
| SaHSP70-23 | CL3978.Contig6_All | 24 | 20 | 5 | 1 | 6 |
| SaHSP70-24 | CL3978.Contig8_All | 25 | 19 | 5 | 1 | 4 |
| SaHSP70-25 | Unigene40279_All | 25 | 24 | 8 | 5 | 3 |
| SaHSP70-26 | Unigene35732_All | 29 | 17 | 6 | 4 | 3 |
| SaHSP70-27 | Unigene45234_All | 27 | 14 | 0 | 0 | 2 |
| SaHSP70-28 | Unigene5923_All | 23 | 16 | 8 | 0 | 3 |
| [1] |
楚宗丽, 李亮杰, 姬虹, 等, 2022. 小麦Hsp70基因家族鉴定及蛋白互作网络分析[J]. 江苏农业科学, 50(10): 37-44 (in Chinese).
|
| [2] |
郝昕, 邓振, 陈洁, 等, 2022. 松材线虫热激蛋白家族成员结构与功能分析[J]. 中国生物防治学报, 38(3): 774-784.
|
| [3] |
何伶俐, 2013. 两种中国南海软珊瑚的化学成分及其生物活性研究[D]. 合肥: 合肥工业大学.
|
| [4] |
黄丁勇, 王建佳, 陈甘霖, 等, 2021. 亚龙湾珊瑚礁大型礁栖生物的群落结构及生态警示[J]. 生态学杂志, 40(2): 412-426.
|
| [5] |
罗洪林, 张瑶瑶, 梁万文, 等, 2015. 罗非鱼HSP70(tHSP70)蛋白的生物信息学分析、真核表达及质谱鉴定[J]. 水产学报, 39(5): 618-627.
|
| [6] |
彭乔烽, 陈朗, 马小梅, 等, 2019. 热休克蛋白HSP70家族功能研究进展[J]. 甘肃畜牧兽医, 49(5): 18-20 (in Chinese).
|
| [7] |
宋刚, 方志刚, 王玉龙, 等, 2022. 柳枝稷Hsp70家族基因鉴定与生物信息学分析[J]. 草业科学, 39(10): 2112-2126.
|
| [8] |
王长云, 刘海燕, 邵长伦, 等, 2008. 软珊瑚(Sinularia flexibilis)和柳珊瑚(Plexaura homomalla)化学防御物质研究进展[J]. 生态学报, 28(5): 2320-2328.
|
| [9] |
王荣青, 万红建, 李志邈, 等, 2014. 番茄Hsp70基因鉴定及系统发育关系分析[J]. 核农学报, 28(3): 378-385.
|
| [10] |
王正, 唐旭利, 李磊, 等, 2018. 西沙短指软珊瑚Sinularia multiflora化学成分研究[J]. 中国海洋药物, 37(5): 47-52.
|
| [11] |
徐秀菊, 2015. 盔形珊瑚属HSP90、Ferritin、CuZnSOD基因的克隆与表达分析[D]. 湛江: 广东海洋大学.
|
| [12] |
张淑红, 范永山, 2022. 玉米大斑病菌热激蛋白Hsp70的鉴定和结构分析[J]. 福建农业学报, 37(9): 1187-1193.
|
| [13] |
AHSEN, PFANNER, 1997. Molecular chaperones: towards a characterization of the heat-shock protein 70 family[J]. Trends in Cell Biology, 7(3): 129-133.
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
NITIKA,
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}