
热带海洋特色生态系统恢复重构与保护思考*
*感谢匿名审稿专家提出的宝贵修改意见和建议。
|
黄良民(1953—), 男, 广东省饶平县人, 研究员, 从事海洋生态学研究。email: hlm@scsio.ac.cn |
Editor: 林强
收稿日期: 2023-12-26
修回日期: 2024-02-07
网络出版日期: 2024-03-13
基金资助
国家自然科学基金项目(42425004)
国家重点研发项目(2022YFC3102400)
国家重点基础研究发展计划(973计划)(2015CB452904)
广东省科技计划项目(2023B1212060047)
Thoughts on the restoration, reconstruction and protection of typical tropical marine ecosystems
Editor: LIN Qiang
Received date: 2023-12-26
Revised date: 2024-02-07
Online published: 2024-03-13
Supported by
National Natural Science Foundation of China(42425004)
National Key Research and Development Program of China(2022YFC3102400)
National Program on Key Basic Research Project (973 Program)(2015CB452904)
Science and Technology Planning Project of Guangdong Province, China(2023B1212060047)
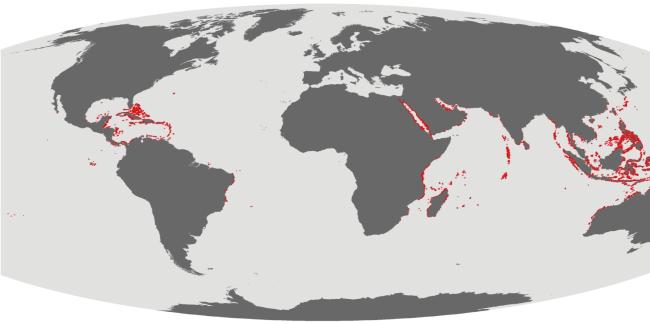
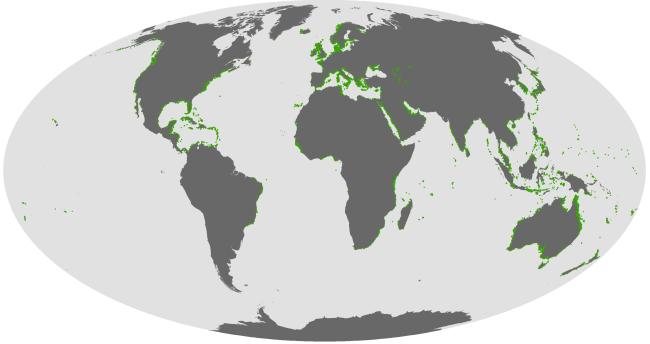
热带海洋特色生态系统是指珊瑚礁、红树林、海草床等由独特生境与生物共同构成的生态系统, 主要分布于全球热带亚热带海区; 其特点是物质循环效率高, 生态功能独特, 具有丰富的生物多样性和极高生产力, 历来是海洋科学研究和资源利用与保护的重点对象。然而, 受人类活动和全球气候变化等多重压力影响, 这些生态系统的退化趋势日益凸显。如何恢复热带海洋特色生态系统的功能, 促进它们的可持续发展并造福人类, 引起了学界和国际社会的高度关注。本文阐述了世界热带海洋特色生态系统的分布格局、退化趋势及其影响因素, 分析了我国珊瑚礁、红树林、海草床生态系统的分布现状和生物多样性特点, 并以珊瑚礁生态系统为重点, 探讨了热带海洋特色生态系统退化的原因, 以及目前我国开展修复和保护的有关行动措施。根据全球生物多样性保护行动计划, 结合我国的实际情况, 提出热带海洋特色生态系统恢复重构与保护的对策建议: 1)建议国家组建南海特色生态系统研究中心、智能监测监控系统、专项数据库和专家智库; 2)设立热带海洋特色生态系统恢复与保护示范区, 组织多学科领域专家联合开展研究—试验—示范—评估—推广—监控; 3)健全政策法规, 加强执法管理, 重视科普教育, 提高公众环保意识, 形成社会自觉行动。这些将为加强热带海洋特色生态系统及其生物多样性的保护和管理, 维护其可持续发展和落实《昆明-蒙特利尔全球生物多样性框架》提出的30×30目标提供参考。
黄良民 , 林强 , 谭烨辉 , 黄小平 , 周林滨 , 黄晖 . 热带海洋特色生态系统恢复重构与保护思考*[J]. 热带海洋学报, 2024 , 43(6) : 1 -12 . DOI: 10.11978/2023200
Typical tropical marine ecosystems are composed of unique habitats and organisms, such as coral reefs, mangroves and seagrass beds. These ecosystems are mainly distributed in tropical and subtropical marine areas worldwide. It is characterized by high material cycling efficiency, unique ecological functions, rich biodiversity and extremely high productivity, and thus a key object of marine scientific research and resource utilization. Under the multiple pressures of human activities and global changes, its degradation trend has become increasingly prominent. How to restore the function of these typical ecosystems and maintain their sustainable development for the benefit of mankind has attracted increasing amounts of attention from the academic community and the international society. In this paper, the distribution patterns, degradation trends and influencing factors of such typical marine ecosystems in the world are described, with an emphasis on the distribution patterns and biodiversity characteristics of coral reef, mangrove and seagrass bed ecosystems in China. Focusing on coral reef ecosystems, this paper discusses the causes of degradation in the ecosystems and relevant action measurements for restoration and protection in China. According to the global action plan for biodiversity conservation and the actual situation of China, countermeasures and suggestions for the restoration, reconstruction and protection of the typical marine ecosystems were proposed: 1) Establishment of a research center for typical ecosystems in the South China Sea, an intelligent monitoring system and database, and an expert think tank; 2) establishing a demonstration zone for the restoration and protection of the typical marine ecosystems, and inviting experts from multiple disciplines to work together on research-experimentation-demonstration-estimate--promotion-monitor; and 3) optimizing policies and regulations, strengthening law enforcement and management, increasing efforts in science popularization and education, raising public awareness of environmental protection, and promoting conscious social actions. This study aims to provide references for strengthening the conservation and management of typical marine ecosystems and biodiversity, maintaining their sustainable development and implementing the 30×30 goals of the Kunming-Montreal Global Biodiversity Framework.
表1 热带海洋特色生态系统的特征、面临威胁与恢复重构路径Tab. 1 Characteristics, threats and restoration and reconstruction paths of typical tropical marine ecosystems |
| 特色生态 系统类型 | 主要特征 | 分布格局 | 影响因素 | 恢复重构路径 | 主要功能 |
|---|---|---|---|---|---|
| 珊瑚礁 | 珊瑚-共生藻体系, 礁栖生物, 小生境, 高效营养循环, 高生物多样性和生产力 | 热带亚热带, 环礁、堡礁、 台礁、沙洲、岸礁 | 人为干扰、热浪, 酸化、生物侵害、 污染物、废弃物等 | 自然恢复+生态修复(有性- 无性繁育, 移植试验, 增殖护养, 示范推广), 监测监控, 保护管理 | 海洋生物多样性 聚集, 岛礁海岸形 成及防护, 食物产出 |
| 红树林 | 耐盐植物, 真红树- 半红树, 净化水质 | 热带亚热带, 沿岸 滩涂、生态交错带 | 人为砍伐、围垦、 外来种入侵等 | 种苗繁育, 人工种植, 生境恢复, 生态修复, 监测监控, 保护管理 | 海岸防护, 生物多 样性维护, 碳存储 |
| 海草床 | 耐盐植物, 底栖 动物, 附生微藻 | 热带—温带, 沿岸滩涂、 潮间带及潮下带 | 暴风雨、围滩 养殖、拖网捕捞、 采挖海产品、污染物等 | 生境维护, 人工繁育, 播种, 栽培, 监测监控, 保护管理 | 固沙护岸, 净化水质, 生物多样性维护, 碳存储 |
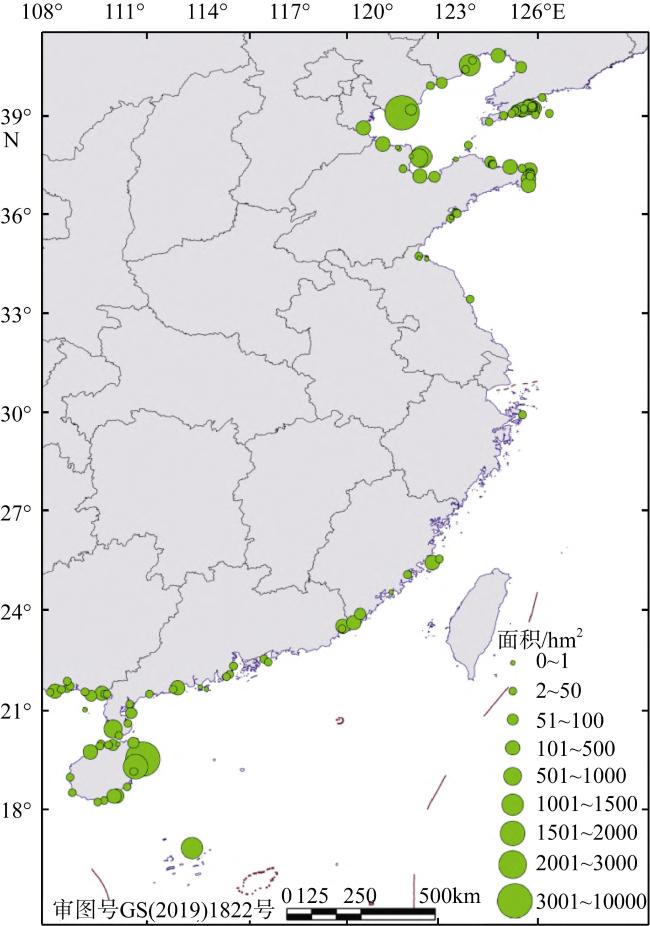
表2 我国红树林面积和真红树种类的分布变化Tab. 2 The distribution variations of mangrove area and true mangrove species in China |
| 分区 | 红树林面积/hm2 | 现存天然种类数 | 已出现灭绝迹象的物种 | 已灭绝的物种 | |
|---|---|---|---|---|---|
| 2001年 | 2019年 | ||||
| 海南 | 3930 | 3666 | 24 | ||
| 广西 | 8375 | 7300 | 11 | 角果木Ceriops tagal | |
| 广东 | 9084 | 14000 | 11 | 角果木Ceriops tagal | |
| 香港 | 510 | 7 | 红海榄Rhizophora stylosa | ||
| 澳门 | 60 | 5 | |||
| 福建 | 615 | 1019 | 5 | 卤蕨Acrostichum aureum, 海漆Excoecaria agallocha | |
| 台湾 | 278 | 6 | 角果木Ceriops tagal | 木榄Bruguiera gymnoihiza | |
| 浙江 | 20 | 33 | |||
| 合计 | 22025* | 26018 | 24 | ||
注: *海南、广西、广东、福建、浙江等地红树林面积来自全国红树林资源专项调查; 合计数字未包括香港、澳门和台湾的红树林面积 |
| [1] |
黄海萍, 陈克亮, 王爱军, 等, 2023. 我国红树林的历史变化、主要问题及保护对策[J]. 海洋开发与管理, 40(2): 125-132.
|
| [2] |
黄晖, 董志军, 练健生, 2008. 论西沙群岛珊瑚礁生态系统自然保护区的建立[J]. 热带地理, 28(6): 540-544.
|
| [3] |
黄晖, 陈竹, 黄林韬, 2021. 中国珊瑚礁状况报告(2010—2019)[M]. 北京: 海洋出版社.
|
| [4] |
黄晖, 俞晓磊, 黄林韬, 等, 2024. 珊瑚礁生态学研究现状和展望[J]. 热带海洋学报, 43(3): 3-12.
|
| [5] |
黄良民, 1991. 南沙群岛海区的光合色素和初级生产力的分布特征初探[M]// 中国科学院南沙综合科学考察队. 南沙群岛及其邻近海区海洋生物研究论文集(2). 北京: 海洋出版社: 34-49.
|
| [6] |
黄良民, 谭烨辉, 宋星宇, 等, 2020. 南沙群岛海区生态过程研究[M]. 北京: 科学出版社.
|
| [7] |
黄小平, 黄良民, 2007. 中国南海海草研究[M]. 广州: 广东经济出版社 (in Chinese).
|
| [8] |
黄小平, 江志坚, 张景平, 等, 2018. 全球海草的中文命名[J]. 海洋学报, 40(4): 127-133.
|
| [9] |
李秀保, 李元超, 许强, 等, 2019. 三亚蜈支洲岛珊瑚礁的现状、生态修复及保护对策[M]. 北京: 科学出版社.
|
| [10] |
廖宝文, 张乔民, 2014. 中国红树林的分布、面积和树种组成[J]. 湿地科学, 12(4): 435-440.
|
| [11] |
全球红树林联盟, 2023. 红树林恢复最佳实践指南(Best practice guidelines for mangrove restoration)[EB/OL]. (2023-10-20) [2023-12-25] https://icriforum.org/guidelines-mangrove-restoration-2023.
|
| [12] |
王友绍, 2013. 红树林生态系统评价与修复技术[M]. 北京: 科学出版社.
|
| [13] |
吴成业, 张建林, 黄良民, 2001. 南沙群岛珊瑚礁潟湖及附近海区春季初级生产力[J]. 热带海洋学报, 20(3): 59-67.
|
| [14] |
余克服, 2012. 南海珊瑚礁及其对全新世环境变化的记录与响应[J]. 中国科学: 地球科学, 42(8): 1160-1172.
|
| [15] |
余克服, 2018. 珊瑚礁科学概论[M]. 北京: 科学出版社: 477-486. YU KEFU, 2018.
Introduction to the science of coral reefs[M]. Beijing: Science Press: 477-486 (in Chinese).
|
| [16] |
赵焕庭, 王丽荣, 袁家义, 2016. 南海诸岛珊瑚礁可持续发展[J]. 热带地理, 36(1): 55-65.
|
| [17] |
郑凤英, 邱广龙, 范航清, 等, 2013. 中国海草的多样性、分布及保护[J]. 生物多样性, 21(5): 517-526.
|
| [18] |
中国科学院南海海洋研究所, 1978. 我国西沙、中沙群岛海域海洋生物调查研究报告集[M]. 北京: 科学出版社 (in Chinese).
|
| [19] |
中国科学院南沙综合科学考察队, 1989. 南沙群岛及其邻近海区综合调查研究报告(一), 上、下卷[R]. 北京: 科学出版社 (in Chinese).
|
| [20] |
中国绿色时报, 2023. 推动红树林保护进入高质量发展阶段—我国红树林保护修复成就综述[EB/OL]. (2023-08-02) [2023-12-15]. http://www.isenlin.cn/sf_ABA0E44EE1F44684AB15B1491CC72A5B_209_A952252C215.html.
|
| [21] |
周毅, 江志坚, 邱广龙, 等, 2023. 中国海草资源分布现状、退化原因与保护对策[J]. 海洋与湖沼, 54(5): 1248-1256.
|
| [22] |
朱袁智, 1981. 西沙群岛岛屿生物礁[J]. 南海海洋科学集刊, (3): 33-48 (in Chinese).
|
| [23] |
邹仁林, 宋善文, 马江虎, 1975. 海南岛浅水造礁石珊瑚[M]. 北京: 科学出版社: 1-65 (in Chinese).
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
IUCN, 2023. Global species action plan: supporting implementation of the Kunming-Montreal global biodiversity framework[R]. Gland, Switzerland: IUCN.
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
UNEP-WCMC,SHORT F T, 2021a. Global distribution of seagrasses (version 7.1). Seventh update to the data layer used in Green and Short (2003)[R]. Cambridge, UK: UN Environment World Conservation Monitoring Centre, Doi: 10.34892/x6r3-d211.
|
| [55] |
UNEP-WCMC,
|
| [56] |
UNEP-WCMC. 2023. Reef futures: Exploring the ecosystem services and potential futures of coral reefs[EB/OL]. (2023-07) [2023-12-15]. https://www.unep-wcmc.org/en/news/reef-futures-exploring-the-ecosystem-services-and-potential-futures-of-coral-reefs.
|
| [57] |
UNESCO, 2023. Great barrier reef[EB/OL]. [2023-12-15]. https://whc.unesco.org/en/list/154.
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}