
南海小叶海蛞蝓(Phyllidiella nanhaiensis sp. nov.)线粒体基因组特征与系统进化
|
刘慧(1999—), 硕士研究生, 主要从事海洋动物行为生理学。email: lh19553590323@s.ytu.edu.cn |
Copy editor: 孙翠慈
收稿日期: 2024-03-01
修回日期: 2024-03-18
网络出版日期: 2024-04-15
基金资助
国家重点研发项目(2022YFC3102403)
国家重点研发项目(2021YFF0502803)
广东省应用海洋生物学重点实验室运行经费(2023B1212060047)
Complete mitogenome data of sea slug Phyllidiella nanhaiensis sp. nov. and its phylogenetic implications
Copy editor: SUN Cuici
Received date: 2024-03-01
Revised date: 2024-03-18
Online published: 2024-04-15
Supported by
National Key Research and Development Program of China(2022YFC3102403)
National Key Research and Development Program of China(2021YFF0502803)
Science and Technology Planning Project of Guangdong Province, China(2023B1212060047)
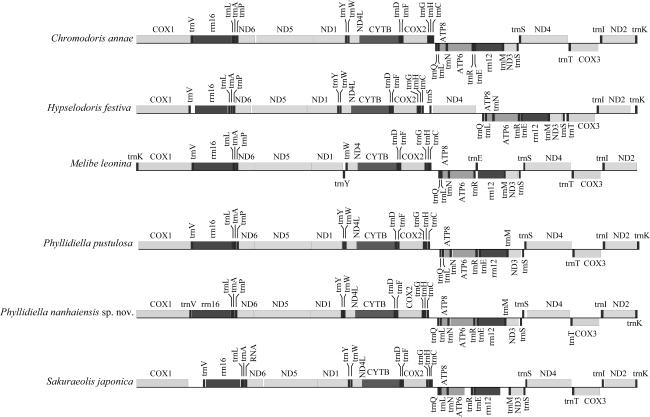
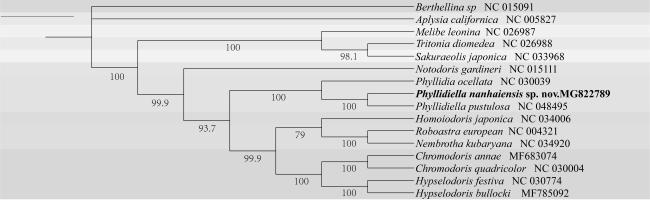
南海小叶海蛞蝓(Phyllidiella nanhaiensis sp. nov.)发现于南海海域, 其形态特征与叶海牛科物种极为相似, 呈修长的卵圆形, 背部呈深灰色且具不规则乳白色瘤状突起, 其中央两排较大橙色突起并行分布。本文利用线粒体全基因组测序对其进行鉴定, 并根据蛋白编码基因(protein-coding genes, PCGs)序列分析其系统进化关系。结果表明, 该物种线粒体基因组全长为14677bp, 包含37个基因, 分别为13个PCGs、2个rRNA和22个tRNA。线粒体基因组的碱基组成为: A占30.4%、T占37.3%、C占15.0%、G占17.3%, A+T含量为67.7%。在13个PCGs中, 除了ND5和ND6使用TTG作为起始密码子外, 其他11个PCGs的起始密码子均符合ATN的形式。最大似然法进化树分析结果表明, 该物种与突丘小叶海蛞蝓(Phyllidiella pustulosa)亲缘关系极为密切, 并与媚眼叶海牛(Phyllidia ocellata)聚为一支。BLAST (basic local alignment search tool)分析发现, 该物种与P. pustulosa线粒体全基因组序列相似度为94.6%, COX1基因序列相似度为98.8%。结合形态特征与分子鉴定结果表明, 该物种为Phyllidiella属新种, 将之命名为南海小叶海蛞蝓(Phyllidiella nanhaiensis sp. nov.)。南海小叶海蛞蝓的发现及其线粒体基因组数据为Phyllidiella属的深入研究提供参考, 对海蛞蝓的物种多样性和系统发育研究具有重要意义。
刘慧 , 张辉贤 , 刘馨蔓 , 林强 , 沈萍萍 . 南海小叶海蛞蝓(Phyllidiella nanhaiensis sp. nov.)线粒体基因组特征与系统进化[J]. 热带海洋学报, 2025 , 44(1) : 1 -8 . DOI: 10.11978/2024049
The sample was obtained in the South China Sea, and its morphological characters are very similar to those of the Phyllidiidae species. It is elongated and ovoid, with a dark grey dorsal surface and irregular creamy-white verrucae, and two rows of orange protrusions in the center of the dorsal surface in parallel. In this study, this species was identified by mitochondrial whole genome sequencing and analyzed based on the sequences of protein-coding genes (PCGs). The results show that the full length of the mitochondrial genome is 14677 bp, containing 37 genes, which are 13 PCGs, 2 rRNAs and 22 tRNAs. The base composition of the mitochondrial genome is 30.4% A, 37.3% T, 15.0% C, and 17.3% G, with a preference for AT (67.7%). Among the 13 PCGs, there are 11 start codons conformed to the codon form of ATN, which used TTG as the start codon. The evolutionary tree was constructed using the maximum likelihood method, and the results show that this species is closely related to Phyllidiella pustulosa and clustered with Phyllidia ocellata. The complete mitochondrial genome sequence similarity between this species and P. pustulosa reached 94.6% by BLAST, and the gene sequence similarity of COX1 reaches 98.8%. Based on combined morphological characters and molecular identification, it is a new species of the genus Phyllidiella in the South China Sea, Phyllidiella nanhaiensis sp. nov.. The mitogenome of P. nanhaiensis sp. nov. will provide a reference for further study of the genus Phyllidiella, which is important for the study of species diversity and phylogeny of sea slugs.
表1 线粒体基因组信息表Tab. 1 List of mitochondrial genome information |
| 物种 | 长度 | GenBank登录号 |
|---|---|---|
| Chromodoris annae | 14260bp | MF683074 |
| Chromodoris quadricolor | 14259bp | NC 030004 |
| Phyllidiella pustulosa | 14717bp | NC 048495 |
| Phyllidia ocellata | 14598bp | NC 030039 |
| Notodoris gardineri | 14424bp | NC 015111 |
| Roboastra europaea | 14472bp | NC 004321 |
| Sakuraeolis japonica | 15059bp | NC 033968 |
| Melibe leonina | 14513bp | NC 026987 |
| Tritonia diomedea | 14540bp | NC 026988 |
| Berthellina sp. | 15688bp | NC 015091 |
| Aplysia californica | 14117bp | NC 005827 |
| Hypselodoris bullocki | 14666bp | MF785092 |
| Hypselodoris festiva | 14880bp | NC 030774 |
| Nembrotha kubaryana | 14395bp | NC 034920 |
| Homoiodoris japonica | 14601bp | NC 034006 |
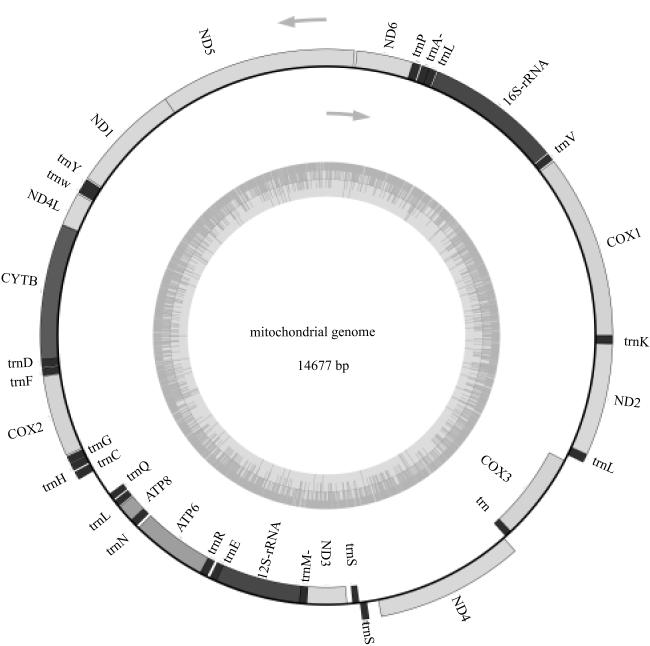
图2 线粒体基因组圈图图中箭头方向表示转录方向, 内圈的灰色部分代表序列的GC含量 Fig. 2 The mitogenome map. The direction of the arrow indicates the direction of transcription. The grey part of the inner ring represents the GC content of the sequence, which is calculated according to a certain window size and can be visualized in the figure |
表2 线粒体基因组的碱基组成Tab. 2 Composition of the mitochondrial genome |
| 基因 | 长度 | A/% | T/% | C/% | G/% | (A+T)/% | AT-skew | GC-skew |
|---|---|---|---|---|---|---|---|---|
| 全基因组 | 14 677 | 30.4 | 37.3 | 15.0 | 17.3 | 67.7 | -0.1023 | 0.0719 |
| 蛋白质编码基因 | 10 777 | 28.8 | 37.8 | 15.5 | 17.9 | 66.6 | -0.1345 | 0.0700 |
| tRNA | 1 379 | 35.0 | 33.1 | 17.3 | 14.7 | 68.1 | 0.0266 | 0.0818 |
| 12S rRNA | 755 | 34.3 | 40.9 | 15.5 | 13.0 | 71.5 | -0.0926 | -0.0884 |
| 16S rRNA | 1 125 | 36.5 | 35.0 | 11.7 | 16.7 | 71.6 | 0.0211 | 0.1750 |
表3 线粒体基因组特征Tab. 3 Characterization of the mitochondrial genome |
| 基因 | 反密码子 | 起始位点 | 终止位点 | 编码链 | 长度 | 起始/终止密码子 |
|---|---|---|---|---|---|---|
| COX1 | / | 1 | 1530 | H | 1530 | ATG/TAA |
| tRNA-Val | TAC | 1542 | 1605 | H | 64 | / |
| 16S rRNA | / | 1614 | 2738 | H | 1125 | / |
| tRNA-Leu | TAG | 2745 | 2811 | H | 67 | / |
| tRNA-Ala | TGC | 2808 | 2875 | H | 68 | / |
| tRNA-Pro | TGG | 2885 | 2950 | H | 66 | / |
| NAD6 | / | 2951 | 3421 | H | 471 | TTG/TAA |
| NAD5 | / | 3438 | 5087 | H | 1650 | TTG/TAG |
| NAD1 | / | 5065 | 5979 | H | 915 | ATG/TAA |
| tRNA-Trp | TCA | 6055 | 6117 | H | 63 | / |
| NAD4L | / | 6127 | 6411 | H | 285 | ATA/TAA |
| CYTB | / | 6413 | 7534 | H | 1122 | ATA/TAA |
| tRNA-Asp | GTC | 7530 | 7599 | H | 70 | / |
| tRNA-Phe | GAA | 7603 | 7667 | H | 65 | / |
| COX2 | / | 7670 | 8347 | H | 678 | ATG/TAG |
| tRNA-Gly | TCC | 8356 | 8419 | H | 64 | / |
| tRNA-His | GTG | 8422 | 8485 | H | 64 | / |
| tRNA-Cys | GCA | 8498 | 8568 | H | 71 | / |
| tRNA-Gln | TTG | 8809 | 8870 | L | 62 | / |
| tRNA-Leu | TAA | 8883 | 8945 | L | 63 | / |
| ATP8 | / | 8953 | 9108 | L | 156 | ATG/TAA |
| tRNA-Asn | GTT | 9110 | 9175 | L | 66 | / |
| ATP6 | / | 9198 | 9866 | L | 669 | ATG/TAA |
| tRNA-Arg | TCG | 9867 | 9933 | L | 67 | / |
| tRNA-Glu | TTC | 9957 | 10024 | L | 68 | / |
| 12S rRNA | / | 10017 | 10771 | L | 755 | / |
| tRNA-Met | CAT | 10769 | 10835 | L | 67 | / |
| NAD3 | / | 10834 | 11187 | L | 354 | ATG/TAA |
| tRNA-Ser | TGA | 11234 | 11293 | L | 60 | / |
| tRNA-Ser | GCT | 11299 | 11359 | H | 61 | / |
| NAD4 | / | 11458 | 12690 | H | 1233 | ATA/TAA |
| tRNA-Thr | TGT | 12710 | 12774 | L | 65 | / |
| COX3 | / | 12770 | 13549 | L | 780 | ATG/TAG |
| tRNA-Ile | GAT | 13604 | 13675 | H | 72 | / |
| NAD2 | / | 13677 | 14610 | H | 934 | ATG/T |
| tRNA-Lys | TTT | 14611 | 14676 | H | 66 | / |
注: PCGs已用加粗字体表示; “/” 表示无数据。Val为缬氨酸, Leu为亮氨酸, Ala为丙氨酸, Pro为脯氨酸, Trp为色氨酸, Asp为天冬氨酸, Phe为苯丙氨酸, Gly为甘氨酸, His为组氨酸, Cys为半胱氨酸, Gln为谷氨酰胺, Asn为天冬酰胺, Arg为精氨酸, Glu为谷氨酸, Met为蛋氨酸, Ser为丝氨酸, Thr为苏氨酸, Ile为异亮氨酸, Lys为赖氨酸 |
| [1] |
李晓露, 2016. 一种海藻Laurencia okamurai和一种软体动物Glossodoris atromarginata的化学成分和生物活性的研究[D]. 上海: 中国科学院上海药物研究所.
|
| [2] |
李忠东, 2015. 神奇的海蛞蝓[J]. 青少年科技博览, (5): 9-10 (in Chinese).
|
| [3] |
林光宇, 张玺, 1965. 海南岛潮间带的后鳃类软体动物[J]. 海洋与湖沼, 7(1): 1-24.
|
| [4] |
刘晓晖, 李晓萌, 李渊, 等, 2020. 南海叶海牛科的研究(异鳃亚纲, 裸鳃目)[J]. 海洋与湖沼, 51(3): 639-643.
|
| [5] |
肖武汉, 张亚平, 2000. 鱼类线粒体DNA的遗传与进化[J]. 水生生物学报, 24(4): 384-391.
|
| [6] |
谢金魁, 曹建国, 1997. 蛞蝓对人肺鳞癌、肺腺癌细胞抑癌作用初探[J]. 肿瘤防治研究, 24(6): 344-346.
|
| [7] |
张丽丽, 程起群, 2012. 鳀科鱼类线粒体全基因组序列结构特征及系统发育信息分析[J]. 海洋渔业, 34(1): 7-14.
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}