
我国海南省东南部台湾那屿藻的形态观察及系统发育分析
|
刘金梅(1996—), 女, 广西玉林市人, 硕士研究生, 主要从事大型海藻的分类研究。 email: |
Copy editor: 林强
收稿日期: 2020-12-24
修回日期: 2021-03-22
网络出版日期: 2021-04-06
基金资助
国家自然科学基金(31670199)
国家自然科学基金(31970216)
天津市教委科研计划项目(JW1705)
版权
Morphological and phylogenetic studies of Yonagunia formosana in southeastern Hainan province, China
Copy editor: LIN Qiang
Received date: 2020-12-24
Revised date: 2021-03-22
Online published: 2021-04-06
Supported by
National Natural Science Foundation of China(31670199)
National Natural Science Foundation of China(31970216)
Scientific Research Plan of Tianjin Municipal Education Committee(JW1705)
Copyright
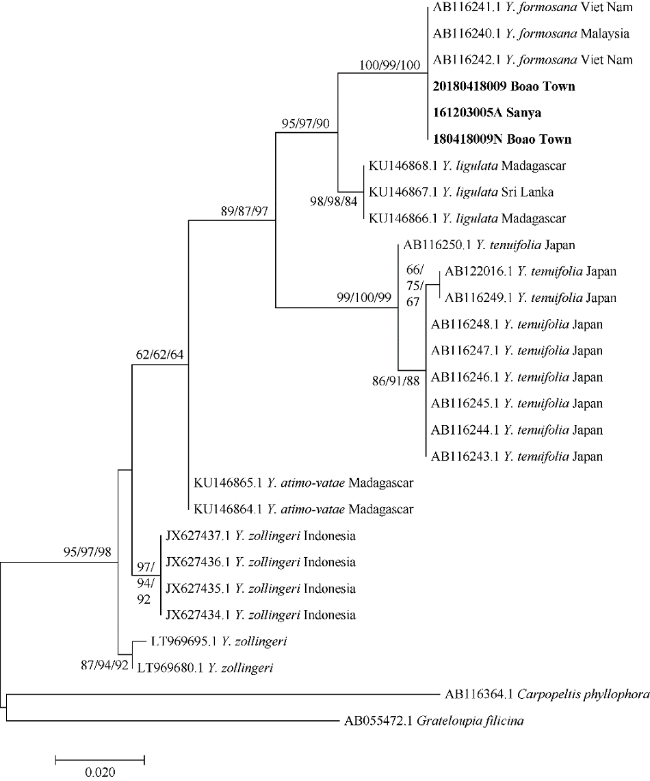
那屿藻属(新拟名)Yonagunia Kawaguchi et Masuda隶属于红藻门海膜科, 本研究通过对该属的台湾那屿藻(新拟名)Yonagunia formosana (Okamura) Kawaguchi et Masuda进行了形态学和基于rbcL序列的系统发育分析, 证实台湾那屿藻存在于我国海南省东南部地区, 并较为详细地描述了该物种的分枝形式、小育枝、皮层细胞等营养结构以及生殖结构, 发现藻体的枝端形状比最初报道的模式种更加多样化, 且藻体各部分结构(分枝形式、小育枝、皮层细胞等)的大小与最初报道的模式种也存在一定的差异。本研究是我国海南省东南部地区台湾那屿藻的首次分子系统发育学研究报道。
刘金梅 , 黄冰心 , 丁兰平 , 王雪聪 , 闫璟 , 闫盼竹 , 张瑶 . 我国海南省东南部台湾那屿藻的形态观察及系统发育分析[J]. 热带海洋学报, 2021 , 40(6) : 76 -82 . DOI: 10.11978/2020150
The genus Yonagunia Kawaguchi et Masuda belongs to Halymeniaceae, Rhodophyta. In this study, we confirmed the existence of Y. formosana in southeastern Hainan Province, China based on the morphological study Yonagunia formosana (Okamura) Kawaguchi et Masuda, combined with the phylogenetic analysis of rbcL sequences. We described in detail the branching form, proliferations, cortical cells, and other vegetative structures and reproductive structures of Y. formosana. We found that the shapes of branches are more diverse, and the size of the structure of the algae, including the branch form, proliferations, and cortical cells and so on, is also different from the type species. This study is the first molecular phylogenetic research report of Y. formosana in southeastern Hainan province.
Key words: Hainan; marine algae; Yonagunia formosana; morphology; rbcL
图1 台湾那屿藻的形态和结构a—b. 藻体外观; c—g.枝端形状; h—i. 藻体小育枝(边缘、表面); j. 藻体部分横切面; k—l. 皮层细胞; m. 髓部横切面; n. 髓部纵切面; o. 小育枝纵切面; p—r. 四分孢子囊; s—t. 果孢子 Fig. 1 Yonagunia formosana: a-b. habit sketch of the frond; c-g. upper portion of the branchlets; h-i. proliferations (edge, surface); j. transection of part of a frond; k-l. cortical cells; m. transection of medulla; n. longitudinal section of medulla; o. longitudinal section of proliferations; p-r. tetrasporangia; s-t. carpospores |
表1 台湾那屿藻、Y. ligulata和样品的比较Tab. 1 Comparison of Y. formosana, Y. ligulata and the sample |
| 结构 | 物种 | ||
|---|---|---|---|
| Y. formosana | Y. ligulata | 本研究样品 | |
| 高度 | 3~8cm | 4~20cm | 2.8~8.5cm |
| 质地 | 软骨质 | 软骨质 | 软骨质 |
| 分枝形式 | 二叉式分枝或近二叉式分枝 | 近二叉式分枝 | 二叉式分枝或不规则分枝 |
| 小育枝 | 表面、边缘均较多, 约5mm | 表面少, 边缘多, 1~15mm | 表面、边缘均较多, 约0.5~3mm |
| 枝端形状 | 枝端钝, 或分叉 | 枝端钝, 或分叉 | 枝端钝, 或较尖, 或分叉, 或着生有小育枝 |
| 皮层 | 60~75μm, 3~7层细胞 | 120~160μm, 4~5层细胞 | 厚约16~83.5μm, 3~9层细胞 |
| 髓部 | 厚250~400μm, 髓丝直径5~10μm | 厚220~400µm, 髓丝直径4~13µm | 厚约260~475μm, 髓丝直径5~13μm |
| 四分孢子囊 | 着生于皮层, 未成熟 | 着生于皮层, 直径9~12μm, 长38~56μm | 着生于皮层, 宽7~9.4μm, 长19~25.6μm |
| 囊果 | 着生于髓部, 球形 | / | 着生于髓部, 近球形 |
| 模式产地 | 中国台湾岛 | 马达加斯加岛 | / |
| 分布地区 | 中国台湾岛和海南岛/马来西亚//越南 | 马达加斯加岛 | 中国海南省三亚/博鳌 |
| 参考文献 | Kawaguchi et al, 2004 | Manghisi et al, 2015 | 本文 |
| [1] |
丁兰平, 黄冰心, 王宏伟, 2015. 中国海洋红藻门新分类系统[J]. 广西科学, 22(2):164-188.
|
| [2] |
黄淑芳, 2000. 台湾东北角海藻图录[M]. 台北, 中国: 国立台湾博物馆: 149.
|
| [3] |
米伟男, 王宏伟, 2009. 海膜科红藻的分子系统学研究进展[C]// 中国生理学会比较生理学专业委员会. “基因、进化与生理功能多样性”海内外学术研讨会暨中国生理学会第七届比较生理学学术会议论文摘要. 大连: 中国生理学会: 1.
|
| [4] |
夏邦美, 2004. 中国海藻志第二卷红藻门第三册[M]. 北京: 科学出版社: 158-160.
|
| [5] |
曾呈奎, 毕列爵, 2005. 藻类名词及名称[M]. 2版. 北京: 科学出版社: 58, 137.
|
| [6] |
赵树雨, 2016. 蜈蚣藻属(Grateloupia C. Agardh)两个新种及亚洲蜈蚣藻个体生态学研究[D]. 辽宁师范大学.
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}