
环境因子对污损生物沙筛贝(Mytilopsis sallei)幼体附着变态的影响研究
|
代丽(1991—), 女, 北京市人, 高级工程师, 博士, 从事核电冷源安全及污损生物防除研究。email: daili0504@163.com |
Copy editor: 林强
收稿日期: 2024-06-13
修回日期: 2024-07-23
网络出版日期: 2024-09-12
基金资助
国家重点研发计划项目(2022YFC3106004)
福建省促进海洋与渔业产业高质量发展专项资助项目(FJHYF-L-2023-14)
福建省海洋服务与渔业高质量发展专项资助项目(FJHY-YYKJ-2023-1-5)
中国核电工程有限公司资助项目(20223160A0857)
Effects of environmental factors on larval settlement and metamorphosis of the fouling organism Mytilopsis sallei
Copy editor: LIN Qiang
Received date: 2024-06-13
Revised date: 2024-07-23
Online published: 2024-09-12
Supported by
National Key Research and Development Program of China(2022YFC3106004)
High-quality Development of Marine and Fishery industry Project of Fujian(FJHYF-L-2023-14)
Ocean Service and High-quality Development of Fishery Project of Fujian(FJHY-YYKJ-2023-1-5)
The Project from China Nuclear Power Engineering Co., Ltd.(20223160A0857)
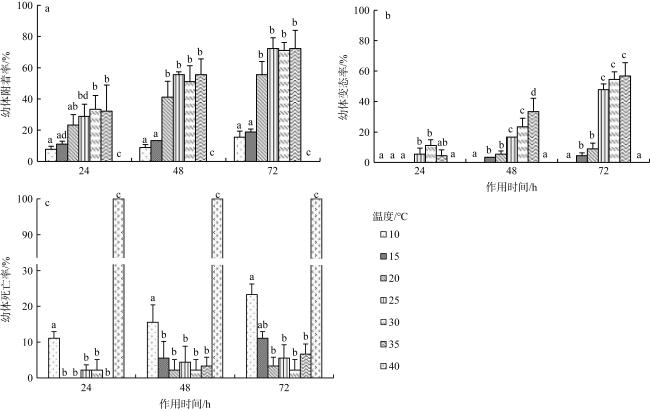
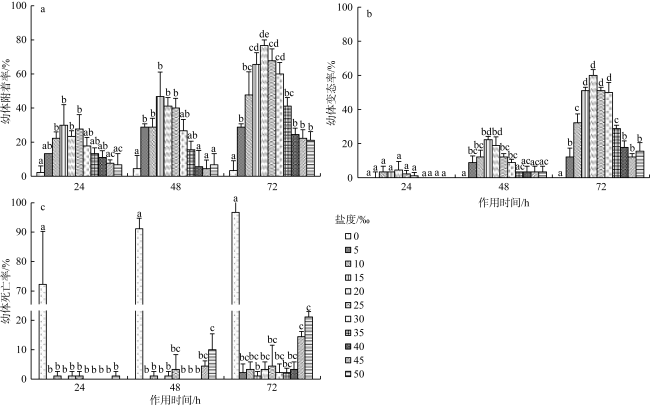
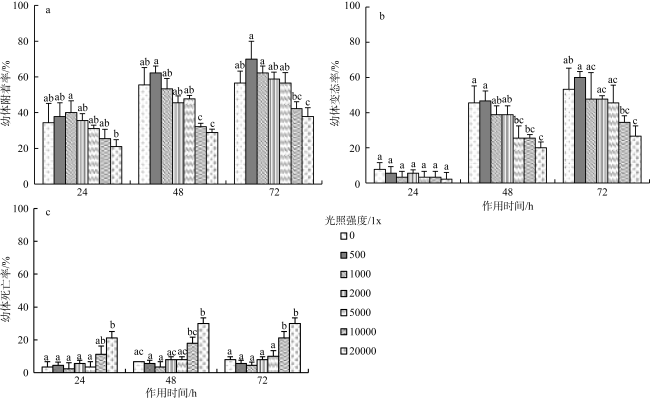
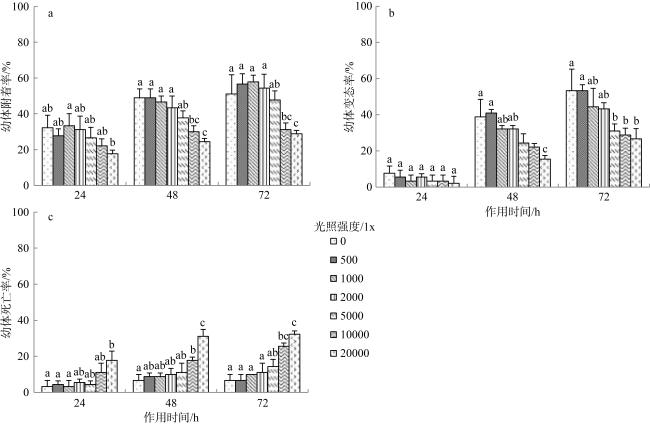
沙筛贝(Mytilopsis sallei)是我国南方一种重要的入侵污损生物, 常见于附着在养殖设施上, 造成渔业损失。本研究通过采用单因子控制试验, 分别检测了不同环境因子(温度、盐度、光照强度和溶解氧)对沙筛贝幼体附着变态的影响。沙筛贝幼体可在25~35℃下正常附着变态, 低温(10℃和15℃)明显抑制了其附着变态, 而高温(40℃)导致幼体全部死亡; 沙筛贝幼体适宜在较广盐度(15‰~30‰)下附着变态, 最适盐度为15‰; 适宜光强范围为0~5000lx, 超过10000lx的光强则会显著降低沙筛贝幼体的附着率和变态率; 沙筛贝幼体具备较强的耐低氧能力, 在40%~100%饱和氧浓度(3.2~8.0mg·L-1)下均能正常附着变态, 但当溶解氧低于20%饱和氧浓度(1.6mg·L-1)时, 则会抑制其附着变态。本研究揭示了沙筛贝幼体附着变态的适宜环境条件, 对预测其在可能分布扩散区域, 并防止其进一步扩张至我国其他海域对当地水产养殖业造成危害具有重要意义。
代丽 , 潘铧康 , 刘励阳 , 何健 , 赵科 , 祁剑飞 , 张真 , 苏培 , 冯丹青 . 环境因子对污损生物沙筛贝(Mytilopsis sallei)幼体附着变态的影响研究[J]. 热带海洋学报, 2025 , 44(3) : 121 -129 . DOI: 10.11978/2024136
The false mussel Mytilopsis sallei is an important invasive fouling organism in the south of China, which often attaches to aquaculture facilities and causes fishery losses. In this study, the ecological single-factor gradient test was used to test larval settlement and metamorphosis of M. sallei under different salinities, water temperatures, light intensities and dissolved oxygen concentrations. The results showed that M. sallei larvae could settle and metamorphose at temperatures between 25℃~35℃. Low temperatures (10℃ and 15℃) significantly inhibited larval settlement and metamorphosis, while high temperature (40℃) caused the death of larvae. M. sallei larvae could settle and metamorphose under a wide salinity range (15‰~30‰), with the optimal salinity of 15‰. The suitable light intensity was 0’5000 lx, but when light intensity was higher than 10000 lx, larval settlement and metamorphosis were significantly inhibited. M. sallei larvae had a strong resistance capacity to low dissolved oxygen concentration. The suitable dissolved oxygen concentration for its settlement and metamorphosis was 40%’100% of saturated oxygen concentration (3.2’8.0 mg·L-1), but when the dissolved oxygen concentration was lower than 20% (1.6 mg·L-1), larval settlement and metamorphosis were inhibited. This study revealed the suitable environmental conditions on larval settlement and metamorphosis of M. sallei, which is important for predicting its possible distribution in the coastal areas of China and preventing its invasion to other areas of China to cause aquaculture industry losses.
图5 沙筛贝幼体在不同光照强度下附着(a)、变态(b)及死亡(c)的情况(光照周期12L:12D)Fig. 5 Larval settlement (a), metamorphosis (b) and death (c) of M. sallei at different light intensities (light cycles 12L:12D) |
| [1] |
蔡立哲, 高阳, 刘炜明, 等, 2006. 外来物种沙筛贝对厦门马銮湾大型底栖动物的影响[J]. 海洋学报, 28(5): 83-89.
|
| [2] |
蒋湘, 魏亦彤, 许乐乐, 等, 2021. 光照强度、温度及海水类型对卤虫(Artemia saline)卵孵化率和幼体存活率的影响[J]. 水产科技情报, 48(4): 197-202.
|
| [3] |
孔宁, 2016. 温度、盐度对皱纹盘鲍“97”选群生长发育的影响[D]. 青岛: 中国科学院研究生院(海洋研究所).
|
| [4] |
马鸿梅, 秦传新, 王兴强, 等, 2020. 应用Maxent建立沙筛贝潜在生境模型[J]. 应用生态学报, 31(4): 1357-1364.
|
| [5] |
牛东红, 王宏蕾, 李家乐, 2024. 海洋贝类对盐度胁迫适应机制的研究进展[J]. 水产学报, 48(4): 049104.
|
| [6] |
王建军, 黄宗国, 郑成兴, 等, 1999. 厦门和东山外来物种沙筛贝的种群动态和结构[J]. 台湾海峡, 18(4): 372-377.
|
| [7] |
张兴志, 何苹萍, 官俊良, 等, 2024. 低氧胁迫及复氧对香港牡蛎抗氧化和能量代谢相关酶活性的影响[J]. 广东海洋大学学报, 44(2): 32-38.
|
| [8] |
张悦, 许道艳, 廖国祥, 等, 2024. 我国海洋外来生物入侵现状、监管问题及建议[J]. 海洋开发与管理, 41(1): 37-44.
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}