
绿华岛4种潮间带大型海藻光合活性对升温及光质的响应
|
区嘉铭(2000—), 男, 广东省珠海市人, 硕士研究生, 从事大型海藻生理生态研究。email: m220501210@st.shou.edu.cn |
Copy editor: 林强
收稿日期: 2024-08-15
修回日期: 2024-09-18
网络出版日期: 2024-09-26
基金资助
南方海洋科学与工程广东省实验室(珠海)资助项目(SML2023SP237)
国家自然科学基金(41876191)
现代农业产业技术体系专项资金(CARS-50)
Response of photosynthetic activity to temperature rise and light quality of four intertidal macroalgae from Lühua Island, Zhejiang, China
Copy editor: LIN Qiang
Received date: 2024-08-15
Revised date: 2024-09-18
Online published: 2024-09-26
Supported by
Southern Marine Science and Engineering Guangdong Laboratory (Zhuhai)(SML2023SP237)
National Natural Science Foundation of China(41876191)
China Agriculture Research System(CARS-50)
区嘉铭 , 汪舒涵 , 赵旭 , 陈健渠 , 孙佳宁 , 邹俏 , 王凯艺 , 章守宇 , 王凯 . 绿华岛4种潮间带大型海藻光合活性对升温及光质的响应[J]. 热带海洋学报, 2025 , 44(3) : 72 -84 . DOI: 10.11978/2024157
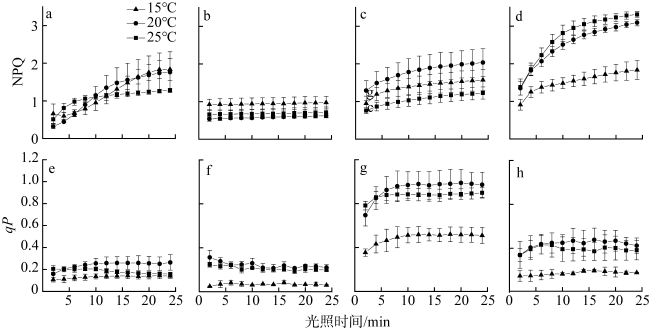
In order to reveal the photosynthetic response of intertidal macroalgae to temperature rise and their photosynthetic efficiency under different light qualities and provide a theoretical basis for the construction of island seaweed farms and the cultivation of seaweed, we used chlorophyll fluorescence and dissolved oxygen sensors to examine the photosynthetic responses of four macroalgae (Ulva pertusa, Grateloupia livida, Sargassum thunbergii and Sargassum fusiforme) from the intertidal zone of Lühua Island to temperatures (15℃, 20℃, and 25℃) and light quality (white, red, green, and blue). The results showed that temperature rise reduced the maximum quantum yield (Fv/Fm) of Photosystem Ⅱ (PSII), but significantly enhanced the actual quantum yield [Y(Ⅱ)], the ratio of photosynthetic repair to damage rate (r/k) and photochemical quenching (qP) under high light intensity (900 μmol photons·m-2·s-1) in these seaweeds. Temperature rise significantly increased the non-photochemical quenching (NPQ) of S. fusiforme, but significantly decreased that of G. livida. Compared to other seaweeds, S. thunbergii exhibited the highest maximum relative electron transfer rate (rETRmax), half-saturated light intensity (Ek), qP and r/k. Under white, red and green light, these seaweeds showed no significant differences in maximum photosynthetic rate (Pmax), but their Pmax was significantly reduced under blue light, with the greatest decrease observed in U. pertusa. As shown in our study, the photosynthetic responses of different species to temperature rise and light quality varied considerably, and short-term warming helps the four intertidal macroalgae to resist strong light at low tide and improve their photosynthetic activity under strong light. Notably, S. thunbergii showed higher photosynthetic activity and better adaptation to strong light and temperature rise than the other three. Additionally, the high photosynthetic efficiency under red and green light reflects their good adaptation to the shallow-water environment in the intertidal zone.
Key words: macroalgae; intertidal zone; temperature rise; light quality; photosynthesis
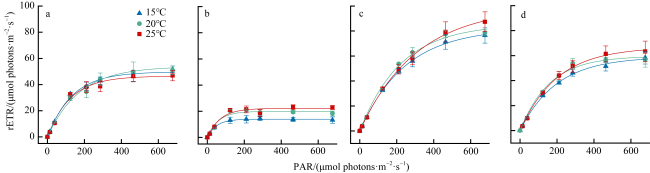
表1 不同温度下孔石莼、舌状蜈蚣藻、鼠尾藻和羊栖菜的Fv/Fm与稳态光曲线拟合参数Tab. 1 Fv/Fm and steady light response curve fitting parameters of U. pertusa, G. livida, S. thunbergii and S. fusiforme at different temperatures |
| 参数 | 温度/℃ | 孔石莼 | 舌状蜈蚣藻 | 鼠尾藻 | 羊栖菜 |
|---|---|---|---|---|---|
| Fv/Fm | 25 | 0.73±0.01b | 0.55±0.04b | 0.71±0.01c | 0.67±0.03b |
| 20 | 0.75±0.01a | 0.55±0.02b | 0.72±0.01b | 0.66±0.01b | |
| 15 | 0.76±0.01a | 0.60±0.02a | 0.74±0.01a | 0.72±0.02a | |
| rETRmax/(μmol electron·m-2·s-1) | 25 | 46.48±1.93b | 22.21 ± 1.14a | 99.04±2.69a | 66.82±1.53a |
| 20 | 53.82±5.20a | 19.93 ± 1.07b | 85.4±3.25b | 59.64±2.29b | |
| 15 | 49.74±1.57ab | 13.98 ± 0.25c | 82.4±1.24c | 59.02±1.33b | |
| α | 25 | 0.38±0.02a | 0.32 ± 0.02b | 0.34±0.00b | 0.33±0.01b |
| 20 | 0.31±0.00b | 0.34 ± 0.03a | 0.38±0.01a | 0.38±0.01a | |
| 15 | 0.38±0.01a | 0.28 ± 0.01c | 0.32±0.00c | 0.30±0.00c | |
| Ek/(μmol photons·m-2·s-1) | 25 | 122.7±8.91b | 69.12±7.36a | 305.85±11.33a | 199.7±6.92a |
| 20 | 172.24±17.47a | 59.05±6.76a | 222.79±12.52c | 158.8±9.86b | |
| 15 | 132.73±7.13b | 49.17±1.90b | 245.46±5.31b | 196.67±6.76a |
注: 表中数据表示为平均值±标准差(mean±SD, n=3), 上标不同字母表示同种海藻的拟合参数在不同温度下的差异性显著(P< 0.05)。 |
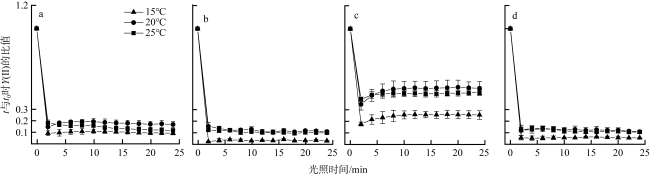
图4 不同温度下4种大型海藻t与t0时刻Y(Ⅱ)的比值随时间的变化(n=3)a. 孔石莼; b. 舌状蜈蚣藻; c. 鼠尾藻; d. 羊栖菜 Fig. 4 Variation of the ratio of Y(Ⅱ) with time at moments t and t0 of four macroalgae at different temperatures (n=3). (a) U. pertusa; (b) G. livida; (c) S. thunbergii; (d) S. fusiforme |
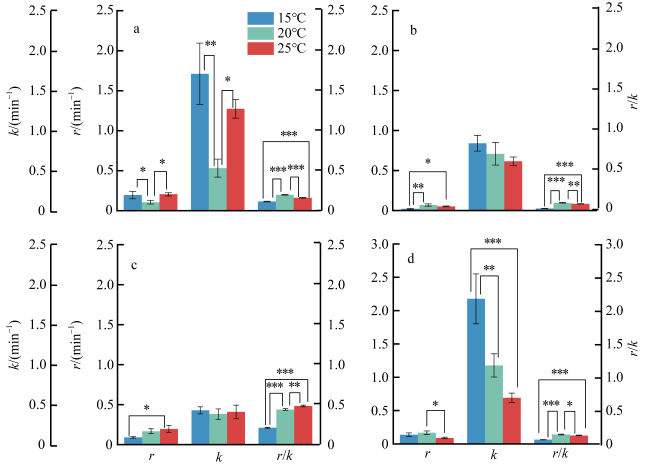
图5 不同温度下4种大型海藻的r、k和r/k值(n=3)a. 孔石莼; b. 舌状蜈蚣藻; c. 鼠尾藻; d. 羊栖菜。*表示显著性差异P<0.05, **P<0.01, ***P<0.001; 无标注则表示无显著差异(P≥0.05) Fig. 5 Values of r, k, and r/k of four macroalgae at different temperatures (n=3). (a) U. pertusa; (b) G. livida; (c) S. thunbergii; (d) S. fusiforme. Values with *, **, and *** denote significant differences of P<0.05, P<0.01, and P<0.001, respectively, and unlabeled values indicate no significant difference (P≥0.05) |
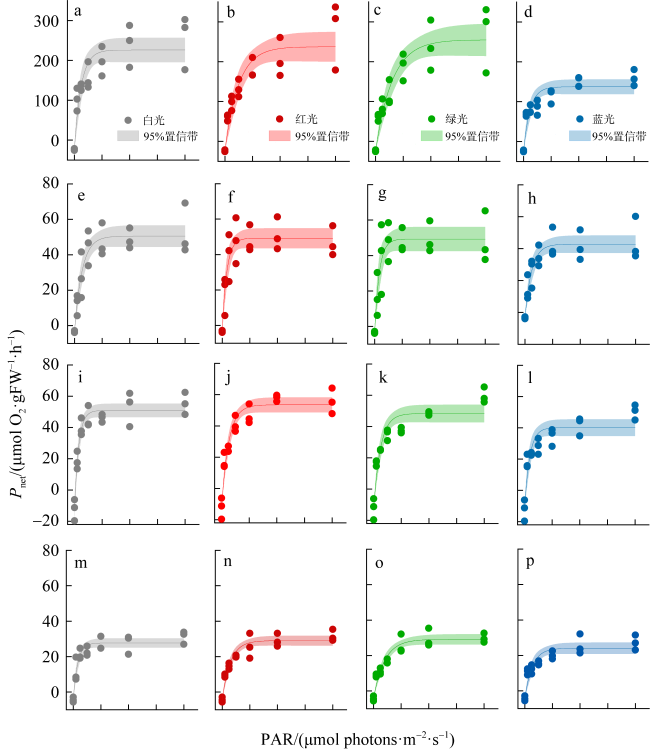
图7 光质对4种大型海藻光合速率的影响(n=3)a—d. 孔石莼; e—h. 舌状蜈蚣藻; i—l. 鼠尾藻; m—p. 羊栖菜 Fig. 7 Effect of light quality on photosynthetic rate of four macroalgae (n=3). (a-d) U. pertusa; (e-h) G. livida; (i-l) S. thunbergii; (m-p) S. fusiforme |
表2 孔石莼、舌状蜈蚣藻、鼠尾藻和羊栖菜的光质P-I曲线拟合参数Tab. 2 Fitting parameters of photosynthesis-irradiance (P-I) curves for U. pertusa, G. livida, S. thunbergii and S. fusiforme under different light qualities |
| 参数 | 光质 | U. pertusa | G. livida | S. thunbergii | S. fusiforme |
|---|---|---|---|---|---|
| Pmax/(μmol O2·g-1FW·h-1) | 白 | 250.28±14.77a | 53.89±2.93a | 63.09±4.26a | 32.05±1.28a |
| 红 | 261.64±17.98a | 52.59±2.71ab | 65.87±2.31a | 33.34±1.34a | |
| 绿 | 279.00±19.68a | 52.70±3.28ab | 60.71±2.70a | 33.44±1.42a | |
| 蓝 | 161.55±8.92b | 46.16±2.64b | 52.49±2.56b | 28.35±1.56b | |
| α(P)/[(μmol O2·g-1FW·h-1)/(μmol photons m-2·s-1)] | 白 | 5.09±1.02a | 0.99±0.18a | 2.09±0.39a | 1.00±0.16a |
| 红 | 2.64±0.48b | 1.59±0.33a | 1.28±0.15b | 0.56±0.07b | |
| 绿 | 2.31±0.40b | 1.55±0.38a | 1.22±0.18b | 0.42±.051b | |
| 蓝 | 3.45±0.66ab | 0.96±0.19a | 1.41±0.26ab | 0.52±0.09b | |
| Rd/(μmol O2·g-1FW·h-1) | — | -24.19±2.00 | -3.38±0.46 | -12.20±3.85 | -4.31±1.16 |
| Ek/(μmol photons m-2·s-1) | 白 | 49.21±11.23b | 54.59±11.11a | 30.22±5.05b | 32.06±5.73b |
| 红 | 98.95±21.49a | 33.13±7.56a | 51.65±6.90a | 59.79±8.78a | |
| 绿 | 120.90±25.50a | 33.96±9.26a | 49.88±8.53a | 79.03±11.26a | |
| 蓝 | 46.84±10.15b | 48.29±10.76a | 37.19±7.71ab | 54.46±11.26ab | |
| Ec/(μmol photons m-2·s-1) | 白 | 5.00±1.00b | 3.53±0.62a | 6.49±1.66b | 4.63±0.74b |
| 红 | 9.60±1.71a | 2.20±0.45a | 10.75±1.24a | 8.28±1.05a | |
| 绿 | 10.96±1.85a | 2.25±0.55a | 11.37±1.69a | 10.91±1.30a | |
| 蓝 | 7.60±1.44ab | 3.67±0.72a | 10.00±1.83ab | 8.99±1.61a |
注: 表中数据表示为平均值±标准差(mean±SD, n=3), 不同字母表示同一物种的拟合参数在不同光质下的差异性显著(P<0.05)。 |
| [1] |
程晓鹏, 2019. 大型海藻光合活性对不同温度和光照的响应[D]. 上海: 上海海洋大学:21.
|
| [2] |
程晓鹏, 章守宇, 林军, 等, 2020. 海带孢子体光合活性对不同温度和光照的响应[J]. 水产学报, 44(2): 234-244.
|
| [3] |
杜响, 骆其君, 陈海敏, 2023. 不同绿藻对模拟酸雨胁迫的生理生化响应特征[J]. 热带海洋学报, 42(5): 115-123.
|
| [4] |
付倩倩, 李航霄, 吴海龙, 等, 2018. 光强对缘管浒苔(Ulva linza)光合生理特性和短期温度效应的影响[J]. 海洋与湖沼, 49(5): 967-974.
|
| [5] |
龚静雨, 2020. 大型海藻生长和光合功能对不同LED光质的响应研究[D]. 广州: 华南理工大学:43.
|
| [6] |
郭赣林, 董双林, 董云伟, 2006. 温度及其波动对孔石莼生长及光合作用的影响[J]. 中国海洋大学学报, 36(6): 941-945.
|
| [7] |
何培民, 段元亮, 刘巧, 等, 2021. 我国近海大型海藻生态修复策略与典型案例[J]. 应用海洋学学报, 40(4): 557-563.
|
| [8] |
黄永健, 崔建军, 陈心怡, 等, 2023. 异枝江蓠对温度和光照强度的光合生理响应[J]. 南方水产科学, 19(4): 139-147.
|
| [9] |
李宝齐, 徐智广, 李凌雪, 等, 2024. 海黍子对强光的光合响应及适应能力研究[J]. 海洋渔业, 46(3): 361-370.
|
| [10] |
李刚, 万明月, 史晓寒, 等, 2022. 中沙大环礁四种大型海藻的光生理特征比较及其对升温的响应[J]. 热带海洋学报, 41(3): 101-110.
|
| [11] |
刘棋琴, 羊芃, 马明婕, 等, 2018. 温度对4种大型海藻氮磷吸收效率及光合生理特性的影响[J]. 水生生物学报, 42(5): 1050-1056.
|
| [12] |
杨宇峰, 罗洪添, 王庆, 等, 2021. 大型海藻规模栽培是增加海洋碳汇和解决近海环境问题的有效途径[J]. 中国科学院院刊, 36(3): 259-269.
|
| [13] |
张建琳, 包炎琳, 孙彬, 等, 2023. 下三横山岛潮间带大型海藻群落构成及季节变化[J]. 应用海洋学学报, 42(2): 225-234.
|
| [14] |
章守宇, 崔潇, 汪振华, 等, 2021. 枸杞岛贻贝养殖筏架附着海藻的群落结构[J]. 水产学报, 45(5): 726-739.
|
| [15] |
赵旭, 2022. 枸杞岛海藻场大型海藻光合生理及生态效应研究[D]. 上海: 上海海洋大学:17.
|
| [16] |
赵旭, 王霄, 李训猛, 等, 2024. 天然海藻场中大型海藻有机碳含量及碳汇能力估算研究——以浙江枸杞岛潮下带为例[J]. 海洋湖沼通报, 46(03): 82-89.
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}