
基于形态和环境DNA宏条形码技术的广西涠洲岛马尾藻场浮游动物群落结构及其粪便碳通量
|
黎洁丽(1999—), 女, 广西壮族自治区玉林市人, 硕士研究生, 从事浮游动物生态研究。email: lijieli@stu2022.jnu.edu.cn |
Copy editor: 林强
收稿日期: 2024-11-15
修回日期: 2024-11-26
网络出版日期: 2024-11-27
基金资助
国家自然科学基金(32271684)
Zooplankton community structure and fecal carbon flux in the seaweed Sargassum beds of Weizhou Island, Guangxi, based on morphological and environmental DNA metabarcoding
Copy editor: LIN Qiang
Received date: 2024-11-15
Revised date: 2024-11-26
Online published: 2024-11-27
Supported by
National Natural Science Foundation of China(32271684)
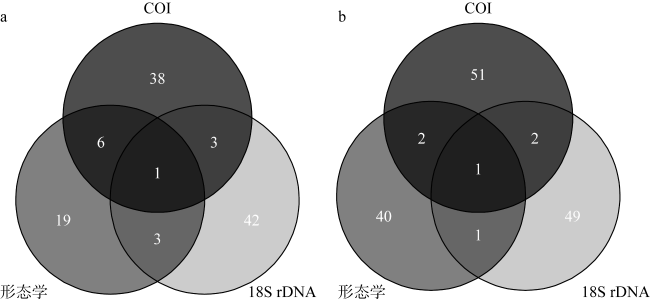
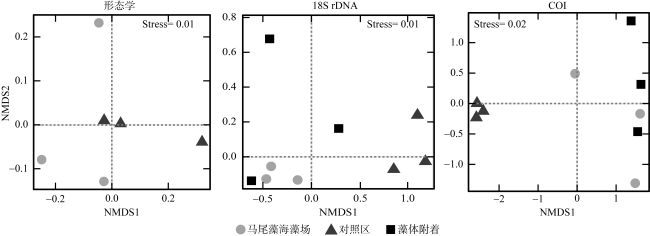
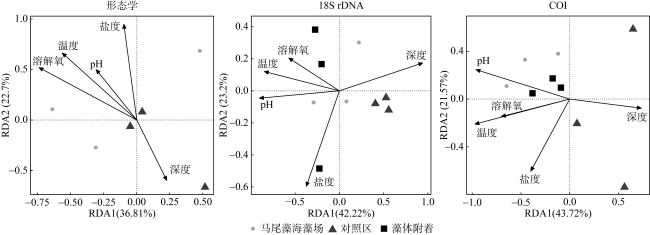
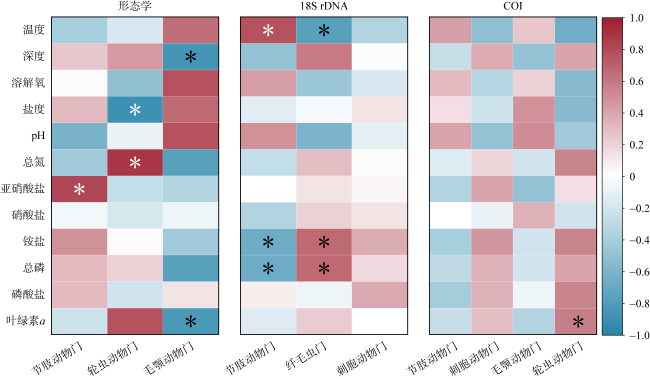
为分析大型海藻对浮游动物群落结构及其粪便碳通量的影响, 于2024年3月、5月和6月在广西涠洲岛马尾藻海藻场及其邻近对照海域进行了浮游动物群落结构调查, 估算了浮游动物粪便碳通量, 并于2024年5月马尾藻茂盛期利用环境 DNA宏条形码对浮游动物群落进行了检测。调查期间基于形态分类学共鉴定浮游动物45种, 其中桡足类38种, 浮游动物丰度变化范围为1160~3960ind.·m-3, 马尾藻海藻场的浮游动物种类及其丰度均高于对照海域(P<0.05)。通过形态学、18S rDNA和COI基因鉴定出的浮游动物种类组成有较大差异, 其中形态学鉴定出较多的桡足类, 18S rDNA检测出较多的纤毛虫, 而 COI基因检测出较多的水母类, 综合三种方法共同检测到的种类为锥形宽水蚤(Temora turbinata)。浮游动物粪便碳通量的变化范围为0.215~0.574mg C·m−2·d−1, 马尾藻海藻场浮游动物的粪便碳通量高于对照海域。调查结果表明, 大型海藻马尾藻场的生态环境能够提高浮游动物的多样性和丰度, 增加浮游动物粪便碳通量。
黎洁丽 , 邹立功 , 杨宇峰 , 王庆 . 基于形态和环境DNA宏条形码技术的广西涠洲岛马尾藻场浮游动物群落结构及其粪便碳通量[J]. 热带海洋学报, 2025 , 44(3) : 104 -120 . DOI: 10.11978/2024207
In order to analyze the effect of seaweed on zooplankton community structure and its fecal carbon flux, a survey of zooplankton community structure was carried out in the Sargassum beds of Weizhou Island and its adjacent waters, Guangxi, in March, May and June 2024, and zooplankton fecal carbon flux was also estimated. When Sargassum bloomed in May 2024, the zooplankton community structure was examined by two types of environmental DNA metabarcoding (18S rDNA and COI gene). A total of 45 zooplankton species, including 38 copepods, were identified by morphology during the survey period. The zooplankton abundance varied from 1160~3960 ind.·m-3, with the zooplankton species and abundance identified in the Sargassum beds being higher than that of its adjacent waters (P<0.05). There were marked differences in the composition of zooplankton species identified by morphology, 18S rDNA and COI gene, with more copepods identified by morphology, more ciliates detected by 18S rDNA, and more medusa species detected by COI gene. The only species detected by all three methods was Temora turbinata. Zooplankton fecal carbon flux varied from 0.215~0.574 mg C·m−2·d−1, and the zooplankton fecal carbon flux in the Sargassum beds were higher than that in its adjacent waters. The results showed that the growth of Sargassum can improve zooplankton diversity and increase zooplankton abundance and fecal carbon flux.
Key words: Weizhou Island; Sargassum beds; zooplankton; environmental DNA; fecal carbon flux
表1 涠洲岛马尾藻海藻场及其邻近海域浮游动物种类组成Tab. 1 Species composition of zooplankton in Sargassum beds of Weizhou Island and its adjacent waters |
| 种类 | 马尾藻海藻场 | 对照区 | 藻体附着 | 18S | COI | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3月 | 5月 | 6月 | 3月 | 5月 | 6月 | 3月 | 5月 | 6月 | |||
| 中华哲水蚤Calanus sinicus | + | + | |||||||||
| 微刺哲水蚤Canthocalanus pauper | + | + | + | + | + | + | |||||
| 小拟哲水蚤Paracalanus parvus | + | + | + | + | + | + | |||||
| 锯缘拟哲水蚤Paracalanus serrulus | + | ||||||||||
| 强额孔雀哲水蚤Parvocalanus crassirostris | + | + | + | + | + | + | |||||
| 瘦拟哲水蚤Paracalanus gracilis | + | + | + | ||||||||
| 矮拟哲水蚤Paracalanus nanus | + | + | |||||||||
| 亚强次真哲水蚤Subeucalanus subcrassus | + | + | + | + | + | + | |||||
| 锥形宽水蚤Temora turbinata | + | + | + | + | + | + | + | + | |||
| 异尾宽水蚤Temora discaudata | + | + | + | ||||||||
| 瘦歪水蚤Tortanus gracilis | + | + | |||||||||
| 太平洋纺锤水蚤Acartia pacifica | + | + | + | + | + | + | |||||
| 红纺锤水蚤Acartia erythraea | + | + | |||||||||
| 瘦尾胸刺水蚤Centropages tenuiremis | + | + | + | + | |||||||
| 奥氏胸刺水蚤Centropages orsinii | + | + | + | + | |||||||
| 椭形长足水蚤Calanopia elliptica | + | + | + | + | + | ||||||
| 长刺小厚壳水蚤Scolecithricella ongispinosa | + | + | |||||||||
| 缘齿厚壳水蚤Scolecithrix nicobarica | + | ||||||||||
| 欧氏后哲水蚤Metacalanus aurivilli | + | + | + | + | |||||||
| 精致真刺水蚤Euchaeta concinna | + | ||||||||||
| 丽隆水蚤Oncaea venusta | + | + | + | + | + | + | + | ||||
| 中隆水蚤Oncaea media | + | + | + | + | + | + | |||||
| 拟长腹剑水蚤Oithona similis | + | + | + | + | + | + | + | ||||
| 坚长腹剑水蚤Oithona rigida | + | + | + | + | + | + | |||||
| 伯氏平头水蚤Candacia bradyi | + | ||||||||||
| 小唇角水蚤Labidocera minuta | + | + | + | ||||||||
| 圆唇角水蚤Labidocera rotunda | + | + | + | + | |||||||
| 叉刺角水蚤Pontella chierchiae | + | + | |||||||||
| 瘦尾筒角水蚤Pontellopsis tenuicauda | + | + | + | ||||||||
| 普通波水蚤Undinula vulgaris | + | + | |||||||||
| 右突新镖水蚤Neodiaptomus schmackeri | + | + | + | + | |||||||
| 奥氏伪镖水蚤Pseudodiaptomus aurivilli | + | + | + | + | |||||||
| 近缘大眼水蚤Corycaeus affinis | + | + | + | + | + | + | |||||
| 小毛猛水蚤Microsetella norvegica | + | + | + | + | + | + | + | + | + | ||
| 红小毛猛水蚤Microsetella rosea | + | + | + | + | + | + | + | + | |||
| 硬鳞暴猛水蚤Clytemnestra scutellata | + | + | + | + | |||||||
| 尖额谐猛水蚤Euterpina acutifrons | + | + | + | + | + | + | + | + | + | ||
| Tisbe sp. | + | + | + | + | + | + | + | + | + | ||
| 疣毛轮虫Synchaeta sp. | + | ||||||||||
| 长尾类幼体Macrura larva | + | + | + | ||||||||
| 端足类Amphipoda sp. | + | + | |||||||||
| 蔓足类幼体Cirripedia larva | + | + | + | + | + | ||||||
| 多毛类幼体Polychaeta larva | + | ||||||||||
| 住囊虫Oikopleura sp. | + | ||||||||||
| 箭虫Sagitta sp. | + | ||||||||||
| 种类合计 | 27 | 37 | 25 | 20 | 25 | 30 | 3 | 5 | 5 | ||
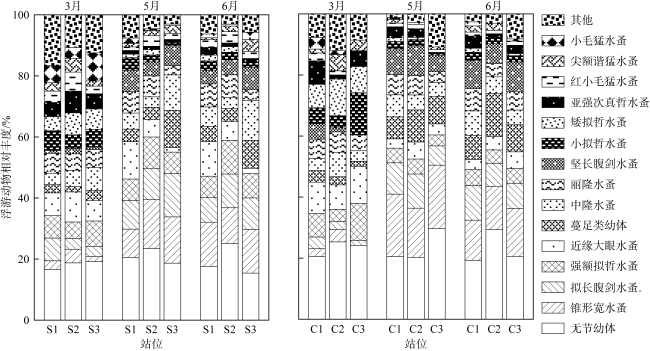
图2 涠洲岛马尾藻场及其邻近海区浮游动物的群落结构a. 不同生境浮游动物种类数; b. 不同月份浮游动物种类数; c. 不同月份和生境的浮游动物丰度; 误差棒上不同字母表示具有显著性差异(P<0.05) Fig. 2 Community structure of zooplankton in Sargassum beds of Weizhou Island and its adjacent waters. (a) Comparison of species number in different habitats; (b) comparison of species number in different months; (c) comparison of zooplankton abundance in different months and habitats. Different letters above the error bars indicate statistically significant differences (P<0.05) |
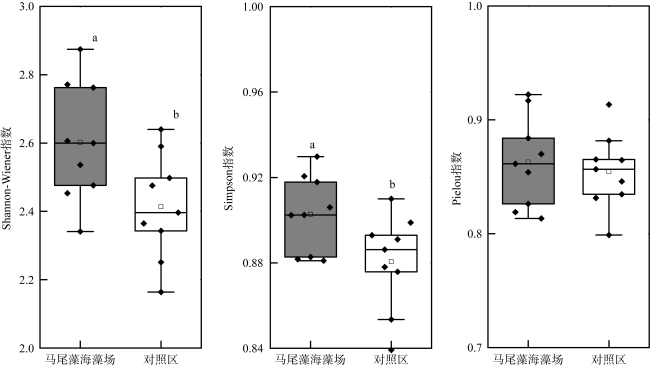
图4 涠洲岛马尾藻场及其邻近海区浮游动物多样性指数Fig. 4 Zooplankton diversity indices in Sargassum beds of Weizhou Island and its adjacent waters |
附表 1 基于形态学、18S rDNA和COI基因的涠洲岛属水平浮游动物组成(S代表马尾藻海藻场, C代表对照区, F代表马尾藻附着样品, 表格统计数据不包括未分类的属)Appendix Tab. 1 The genus composition of zooplankton off Weizhou Island based on morphology, 18S rDNA and COI gene analyses (S for Sargassum beds, C for its adjacent control waters, F for Sargassum attached samples ; unclassified genera are excluded from statistical calculations in this table) |
| 类群 | 属 | 形态学 | 18S rDNA | COI | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| S | C | F | S | C | F | S | C | F | ||
| 桡足类 | 哲水蚤属Calanus | + | + | + | ||||||
| 刺哲水蚤属Canthocalanus | + | + | + | + | ||||||
| 拟哲水蚤属Paracalanus | + | + | + | + | + | |||||
| 隆哲水蚤属Acrocalanus | + | |||||||||
| 基齿哲水蚤属Clausocalanus | + | |||||||||
| 孔雀哲水蚤属Parvocalanus | + | + | + | |||||||
| 次真哲水蚤属Subeucalanus | + | + | + | |||||||
| 宽水蚤属Temora | + | + | + | + | + | + | + | + | ||
| 歪水蚤属Tortanus | + | |||||||||
| 桡足类 | 纺锤水蚤属Acartia | + | + | |||||||
| 胸刺水蚤属Centropages | + | + | + | |||||||
| 长足水蚤属Calanopia | + | + | ||||||||
| 厚壳水蚤属Scolecithrix | + | |||||||||
| 后哲水蚤属Metacalanus | + | |||||||||
| 真刺水蚤属Euchaeta | + | |||||||||
| 隆水蚤属Oncaea | + | + | ||||||||
| Haplostoma | + | |||||||||
| Critomolgus | + | + | ||||||||
| Scambicornus | + | |||||||||
| 中剑水蚤属Mesocyclops | + | |||||||||
| 长腹剑水蚤属Oithona | + | + | + | + | + | + | ||||
| 平头水蚤属Candacia | + | |||||||||
| 唇角水蚤属Labidocera | + | + | ||||||||
| 筒角水蚤属Pontellopsis | + | + | ||||||||
| 波水蚤属Undinula | + | + | ||||||||
| 新镖水蚤属Neodiaptomus | + | + | ||||||||
| 伪镖水蚤属Pseudodiaptomus | + | + | + | |||||||
| 大眼水蚤属Corycaeus | + | + | ||||||||
| 小毛猛水蚤属Microsetella | + | + | + | |||||||
| Andromastax | + | + | ||||||||
| Filexilia | ||||||||||
| 拟双倍猛水蚤属Paramphiascella | + | |||||||||
| 盔头猛水蚤属Clytemnestra | + | |||||||||
| Sentiropsis | + | + | + | |||||||
| 谐猛水蚤属Euterpina | + | + | + | |||||||
| 日角猛水蚤属Tisbe | + | + | + | |||||||
| 盗鳋属Bomolochus | + | |||||||||
| Clausia | + | |||||||||
| Lichomolgus | + | |||||||||
| Pseudanthessius | + | |||||||||
| Asterocheres | + | |||||||||
| Ditrichocorycaeus | + | + | + | |||||||
| 疮痂鱼虱属Lepeophtheirus | + | + | ||||||||
| 轮虫 | 疣毛轮属Synchaeta | + | ||||||||
| 晶囊轮属Asplanchna | + | |||||||||
| 端足类 | 藻钩虾属Ampithoe | + | + | |||||||
| Echinogammarus | + | |||||||||
| Niphargus | + | + | ||||||||
| Paraphronima | + | |||||||||
| Awacaris | + | |||||||||
| Scina | + | |||||||||
| Uristes | + | |||||||||
| 水母类 | Gyractis | + | ||||||||
| 枝手水母属Cladonema | + | |||||||||
| Stauridiosarsia | + | + | ||||||||
| 感棒水母属Laodicea | + | + | + | + | ||||||
| 深杯水母属Abylopsis | + | + | ||||||||
| Sminthea | + | |||||||||
| 棍螅属Coryne | + | |||||||||
| 长管水母属Sarsia | + | |||||||||
| Slabberia | + | |||||||||
| 拟面具水母属Pandeopsis | + | + | ||||||||
| 枝管水母属Proboscidactyla | + | |||||||||
| 多管水母属Aequorea | + | + | ||||||||
| 指突水母属Blackfordia | + | |||||||||
| Bonneviella | + | + | ||||||||
| 钟螅属Campanularia | + | |||||||||
| 直杯螅属Orthopyxis | + | |||||||||
| 钩手水母属Gonionemus | + | |||||||||
| 盛装水母属Agalma | + | |||||||||
| Marrus | + | |||||||||
| 离翼水母属Apolemia | + | |||||||||
| 双生水母属Diphyes | + | |||||||||
| 海蜇属Rhopilema | + | |||||||||
| 被囊类 | 住囊虫属Oikopleura | + | ||||||||
| 毛颚类 | 箭虫属Sagitta | + | + | |||||||
| 纤毛虫 | Durchoniella | + | ||||||||
| Uronemella | + | |||||||||
| Aspidisca | + | |||||||||
| Hemigastrostyla | + | |||||||||
| Amphisiella | + | |||||||||
| 拟铃虫属Tintinnopsis | + | |||||||||
| Coxliella | + | |||||||||
| Amphorellopsis | + | |||||||||
| Eutintinnus | + | |||||||||
| 网纹虫属Favella | + | + | + | |||||||
| Holosticha | + | |||||||||
| Apokeronopsis | + | + | ||||||||
| Thigmokeronopsis | + | |||||||||
| Trichototaxis | ||||||||||
| Cyrtostrombidium | + | |||||||||
| Laboea | + | + | ||||||||
| Parallelostrombidium | + | + | ||||||||
| 旋游虫属Spirostrombidium | + | + | ||||||||
| 急游虫属Strombidium | + | |||||||||
| 旋曳尾虫属Spirotontonia | + | + | ||||||||
| [1] |
陈清潮, 章淑珍, 1974a. 南海的浮游桡足类Ⅰ.[J]. 海洋科学集刊, (9): 101-116.
|
| [2] |
陈清潮, 沈嘉瑞, 1974b. 南海的浮游桡足类Ⅱ.[J]. 海洋科学集刊, (9): 126-137.
|
| [3] |
郭键林, 孙显, 杨宇峰, 等, 2023. 广西涠洲岛大型海藻场及其邻近海域桡足类群落结构演替特征[J]. 热带海洋学报, 42(4): 155-165.
|
| [4] |
何培民, 刘媛媛, 张建伟, 等, 2015. 大型海藻碳汇效应研究进展[J]. 中国水产科学, 22(3): 588-595.
|
| [5] |
黄邦钦, 柳欣, 2015. 边缘海浮游生态系统对生物泵的调控作用[J]. 地球科学进展, 30(3): 385-395.
|
| [6] |
蒋日进, 章守宇, 毕远新, 等, 2015. 枸杞岛海藻场小型无脊椎动物的食物来源[J]. 水产学报, 39(10): 1487-1498.
|
| [7] |
康子清, 张银龙, 吴永波, 等, 2022. 环境DNA宏条形码在生物多样性研究与监测中的应用[J]. 生物技术通报, 38(1): 299-310.
|
| [8] |
李开枝, 尹健强, 黄良民, 等, 2005. 珠江口浮游动物的群落动态及数量变化[J]. 热带海洋学报, 24(5): 60-68.
|
| [9] |
连光山, 王彦国, 孙柔鑫, 等, 2018. 中国海洋浮游桡足类多样性[M]. 北京: 海洋出版社: 40-747.
|
| [10] |
杨宇峰, 罗洪添, 王庆, 等, 2021. 大型海藻规模栽培是增加海洋碳汇和解决近海环境问题的有效途径[J]. 中国科学院院刊, 36(3): 259-269.
|
| [11] |
尹健强, 黄良民, 李开枝, 等, 2013. 南海西北部陆架区沿岸流和上升流对中华哲水蚤分布的影响[J]. 海洋学报, 35(2): 143-153.
|
| [12] |
尹天齐, 王庆, 杨宇峰, 等, 2022. 基于形态学和DNA分子鉴定的珠江口浮游动物群落结构比较研究[J]. 热带海洋学报, 41(3): 172-185.
|
| [13] |
章守宇, 刘书荣, 周曦杰, 等, 2019. 大型海藻生境的生态功能及其在海洋牧场应用中的探讨[J]. 水产学报, 43(9): 2004-2014.
|
| [14] |
张武昌, 陶振铖, 赵苑, 等, 2019. 中国海浮游桡足类图谱[M]. 2版. 北京: 科学出版社.
|
| [15] |
左涛, 李永涛, 左明, 等, 2022. 黄河口邻近海域浮游幼虫群落结构季节变化[J]. 海洋学报, 44(4): 47-56.
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
/
| 〈 |
|
〉 |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}