
绿华岛海域海藻场大型底栖生物群落结构及其对环境因子的响应
|
孙佳宁(2000—), 女, 山东省潍坊市人, 硕士研究生, 从事大型底栖生物生态研究。email: m220501187@st.shou.edu.cn |
Copy editor: 孙翠慈
收稿日期: 2024-10-08
修回日期: 2024-11-05
网络出版日期: 2024-12-16
基金资助
国家重点研发计划项目(2018YFD0900904)
国家重点研发计划项目(2019YFD0901303)
国家自然科学基金(41876191)
Structure of macrobenthic community in the seaweed beds of Lvhua Island waters and its response to environmental factors
Received date: 2024-10-08
Revised date: 2024-11-05
Online published: 2024-12-16
Supported by
National Key Research and Development Program of China(2018YFD0900904)
National Key Research and Development Program of China(2019YFD0901303)
National Natural Science Foundation of China(41876191)
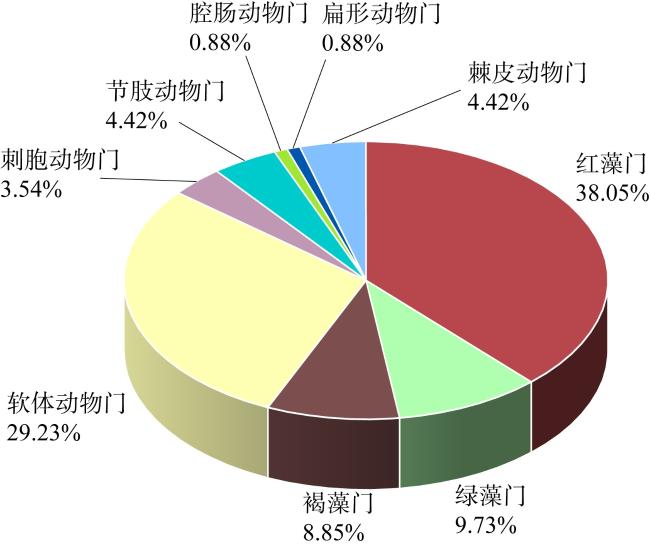
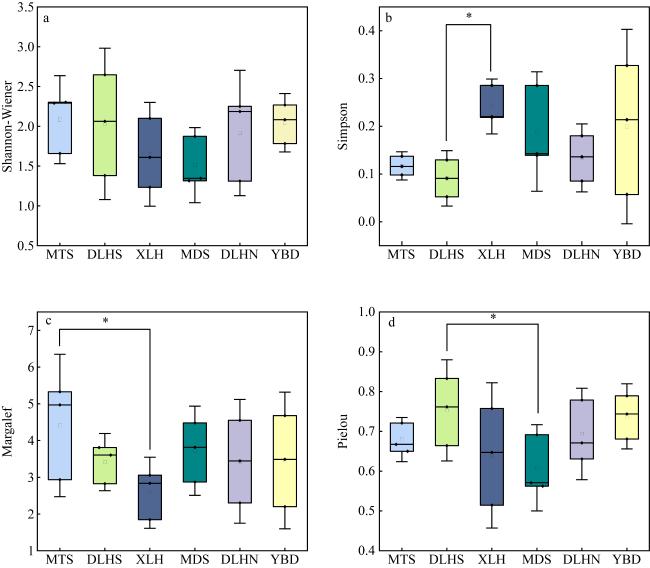
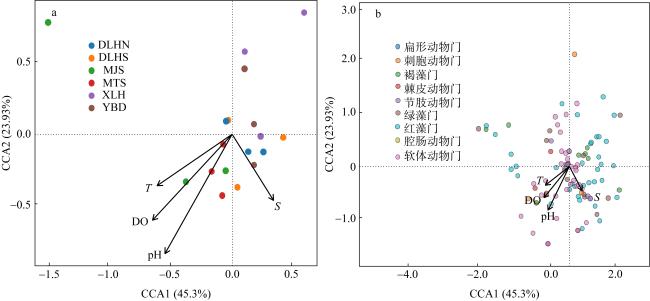
为深入探讨影响绿华岛海域大型海藻及底栖动物种群动态变化的关键因素, 本研究通过现场生态调查, 采用相对重要性指数 (index of relative importance, IRI)、生物多样性指数及典范对应分析(canonical correspondence analysis, CCA)等方法, 结合海藻场大型海藻的生长时期, 即幼苗期(2021年8—9月)、生长期(11—12月)和茂盛期(2022年5—6月), 分析了绿华岛及其邻近海域的大型底栖生物群落组成、生物多样性及与种群变动相关的环境因子。研究结果表明: (1)绿华岛及邻近海域海藻场共鉴定出大型底栖生物113种, 包括49种底栖动物和64种大型海藻, 分属九大门类。其中软体动物门和红藻门的种类数最多, 红藻门在各生长时期均表现出种类优势。瓦氏马尾藻(Sargassum vachellianum)、铜藻(Sargassum horneri)、锈凹螺(Chlorostoma rustica)和紫海胆(Anthocidaris crassispina)为主要优势种。九大门类在三个生长时期的平均丰度为30ind·m-2, 平均生物量为23.83g·m-2。(2)不同生长时期的生物物种数有所差异, 幼苗期最高。丰度与生物量的变动与水温的周期性变化密切相关, 丰度表现为幼苗期(36ind·m-2)>茂盛期(30ind·m-2)>生长期(23ind·m-2), 生物量变化也为幼苗期(25.40g·m-2)>茂盛期(23.88g·m-2)>生长期(22.20g·m-2)。主坐标分析(principal coordinates analysis, PCoA)显示, 不同时期大型底栖生物群落结构存在显著差异(P<0.05)。(3)鳗头山站点的底栖生物种类、丰度、生物量及生物多样性均为最高, 鳗对山站点的多样性最低, 西绿华站点的物种数最少。东绿华南站点的优势种和重要种差异较小, 生物量和丰度处于最低水平; 而贻贝养殖区站点各时期优势种和重要种的重合度较低。(4)除盐度外, 水温(P=0.009)、pH(P=0.001)和溶解氧(P=0.002)均为影响绿华岛海域大型底栖生物分布的重要环境因子, 而盐度(P=0.149)的影响不显著。不同站点的生物对环境因子的适应性存在差异, 鳗头山和鳗对山站点的群落结构与4个环境因子呈正相关, 而贻贝地、西绿华和东绿华北站点则呈负相关。在九个门类中, 褐藻门与4个环境因子呈负相关, 而盐度对腔肠动物门有一定的正相关关系。本研究有助于加深对大型底栖生物在海藻场生态系统中调控作用的认知, 同时也为研究区域生物资源的保护与利用、海藻场建设及生态修复提供科学依据。
孙佳宁 , 王玉清 , 章守宇 , 王凯 . 绿华岛海域海藻场大型底栖生物群落结构及其对环境因子的响应[J]. 热带海洋学报, 2025 , 44(4) : 119 -135 . DOI: 10.11978/2024188
To explore the factors influencing the population dynamics of macroalgae and benthic fauna in the marine area surrounding Lvhua Island, this study conducted field ecological surveys employing the index of relative importance (IRI), biodiversity indices, and canonical correspondence analysis (CCA). These methods were applied across three macroalgal growth stages in the algal beds: seedling (August-September 2021), growth (November-December 2021), and flourishing (May-June 2022), to analyze the community composition, biodiversity, and environmental factors associated to macrobenthic population changes in Lvhua Island and its adjacent waters. The results revealed the following: (1) A total of 113 macrobenthic species were identified in the algal beds of Lvhua Island and its neighboring waters, including 49 species of benthic fauna and 64 macroalgal species, belonging to nine phyla. Among them, Mollusca and Rhodophyta had the highest number of species, with Rhodophyta dominating species diversity across all observed periods. The dominant species included Sargassum vachellianum, Sargassum horneri, Chlorostoma rustica, and Anthocidaris crassispina. The average abundance of the nine phyla across the three growth stages was 30 ind·m-2, and the average biomass was 23.83 g·m-2. (2) Species richness varied by growth stage, peaking during the seedling stage. Abundance and biomass fluctuations were closely tied to periodic water temperature changes, with abundance ranked as seedling stage (36 ind·m-2) > flourishing stage (30 ind·m-2) > growth stage (23 ind·m-2), and biomass following a similar trend: seedling stage (25.40 g·m-2) > flourishing stage (23.88 g·m-2) > growth stage (22.20 g·m-2). Principal coordinates analysis (PCoA) indicated significant differences in macrobenthic community structure across different growth stages (P<0.05). (3) The Mentoushan station recorded the highest species richness, abundance, biomass, and biodiversity, while Menduishan station exhibited the lowest diversity, and the Xilvhua station had the fewest species. Dominant and important species showed minimal variation at Donglvhua South station, where both biomass and abundance were lowest, whereas the mussel farming area displayed relatively low overlap between dominant and important species. (4) Except for salinity, water temperature (P=0.009), pH (P=0.001), and dissolved oxygen (P=0.002) significantly influenced macrobenthic distribution in Lvhua Island waters, while salinity (P=0.149) had no significant effect. Organismal adaptability to environmental factors varied by station: Mentoushan and Menduishan stations showed positive correlations with all four environmental factors, whereas Yibeidi, Xilvhua, and Donglvhua North stations exhibited negative correlations. Among the nine phyla, Phaeophyta correlated negatively with all environmental factors, while Cnidaria showed a positive correlation with salinity. This study enhances understanding of the regulatory role of macrobenthic organisms in algal bed ecosystems and provides a scientific foundation for biological resource conservation, algal bed construction, and ecological restoration in the study area.
Key words: Lvhua Island; seaweed beds; macroalgae; macrozoobenthos; growth stages; environmental factors
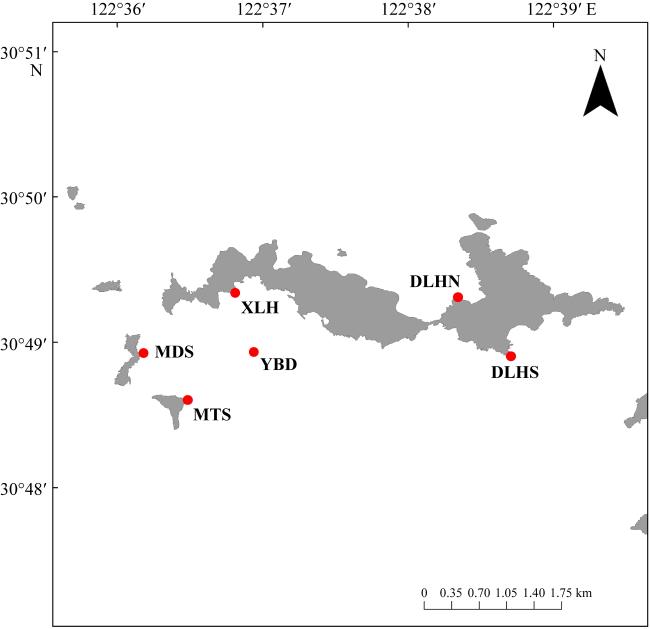
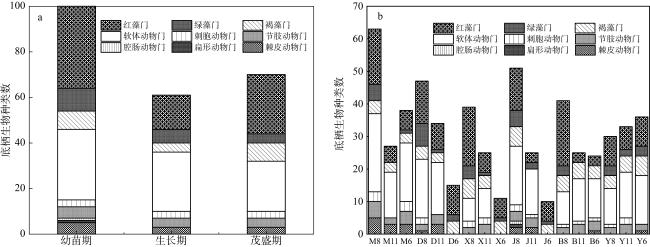
图3 不同生长时期及不同站点大型底栖生物种类组成图3b中M、D、X、J、B、Y分别代表MTS、DLHS、XLH、MDS、DLHN、YBD六个采样站点; 8、11、6分别代表幼苗期、生长期、茂盛期三个时期 Fig. 3 Composition of macrozoobenthos species at different growth stages and sampling stations. In Fig. 3b, the letters M, D, X, J, B, and Y represent the six sampling stations: MTS, DLHS, XLH, MDS, DLHN, and YBD, respectively. The numbers 8, 11, and 6 correspond to the three growth stages: seedling stage, growth stage, and flourishing stage |
表1 海藻场大型底栖生物优势种时空分布Tab. 1 Spatiotemporal distribution of dominant macrobenthic species in seaweed beds |
| 站点 | 时期 | 优势种和重要种 |
|---|---|---|
| MTS | 幼苗期 | **瓦氏马尾藻 Sargassum vachellianum、羊栖菜 Sargassum fusiforme、锈凹螺Chlorostoma rustica、**紫海胆 Anthocidaris crassispina、黄口荔枝螺 Thais luteostoma、甲虫螺 Cantharus cecillei |
| 生长期 | **瓦氏马尾藻 Sargassum vachellianum、珊瑚藻 Corallina officinalis、锈凹螺 Chlorostoma rusticum、**紫海胆 Anthocidaris crassispina、黄口荔枝螺 Thais luteostoma | |
| 茂盛期 | **瓦氏马尾藻 Sargassum vachellianum、羊栖菜 Sargassum fusiforme、锈凹螺 Chlorostoma rusticum、**紫海胆 Anthocidaris crassispina、黄口荔枝螺 Thais luteostoma、甲虫螺 Cantharus cecillei | |
| DLHS | 幼苗期 | **铜藻 Sargassum horneri、鼠尾藻 Sargassum thunbergii、**锈凹螺 Chlorostoma rusticum、**紫海胆 Anthocidaris crassispina、黄口荔枝螺 Thais luteostoma、甲虫螺 Cantharus cecillei |
| 生长期 | 铜藻 Sargassum horneri、**珊瑚藻 Corallina officinalis、**锈凹螺 Chlorostoma rusticum、 黄口荔枝螺 Thais luteostoma、甲虫螺 Cantharus cecillei | |
| 茂盛期 | **铜藻 Sargassum horneri、羊栖菜 Sargassum fusiforme、**锈凹螺 Chlorostoma rusticum、 黄口荔枝螺 Thais luteostoma、甲虫螺 Cantharus cecillei | |
| XLH | 幼苗期 | **瓦氏马尾藻 Sargassum vachellianum、**锈凹螺 Chlorostoma rusticum、黄口荔枝螺 Thais luteostoma、甲虫螺 Cantharus cecillei |
| 生长期 | **瓦氏马尾藻 Sargassum vachellianum、羊栖菜 Sargassum fusiforme、锈凹螺 Chlorostoma rusticum、黄口荔枝螺 Thais luteostoma、甲虫螺 Cantharus cecillei | |
| 茂盛期 | **瓦氏马尾藻 Sargassum vachellianum、囊藻 Colpomenia sinuosa、**锈凹螺 Chlorostoma rusticum、黄口荔枝螺 Thais luteostoma、甲虫螺 Cantharus cecillei | |
| MDS | 幼苗期 | **瓦氏马尾藻 Sargassum vachellianum、羊栖菜 Sargassum fusiforme、**锈凹螺 Chlorostoma rusticum、**紫海胆 Anthocidaris crassispina、黄口荔枝螺 Thais luteostoma、甲虫螺 Cantharus cecillei |
| 生长期 | **瓦氏马尾藻Sargassum vachellianum、 珊瑚藻Corallina officinalis、**锈凹螺 Chlorostoma rusticum、**紫海胆 Anthocidaris crassispina、黄口荔枝螺 Thais luteostoma | |
| 茂盛期 | **瓦氏马尾藻 Sargassum vachellianum、羊栖菜 Sargassum fusiforme、**锈凹螺Chlorostoma rusticum、**紫海胆 Anthocidaris crassispina、 黄口荔枝螺 Thais luteostoma、甲虫螺 Cantharus cecillei | |
| DLHN | 幼苗期 | **羊栖菜 Sargassum fusiforme、鼠尾藻 Sargassum thunbergii、**锈凹螺 Chlorostoma rusticum、甲虫螺 Cantharus cecillei |
| 生长期 | **羊栖菜 Sargassum fusiforme、鼠尾藻 Sargassum thunbergii、**锈凹螺 Chlorostoma rusticum、甲虫螺 Cantharus cecillei、疣荔枝螺 Reishia clavigera | |
| 茂盛期 | **羊栖菜 Sargassum fusiforme、鼠尾藻 Sargassum thunbergii、**锈凹螺 Chlorostoma rusticum、甲虫螺 Cantharus cecillei、疣荔枝螺 Reishia clavigera | |
| YBD | 幼苗期 | 石莼 Ulva lactuca L.、铜藻 Sargassum horneri、舌状蜈蚣藻 Grateloupia livida、瘤荔枝螺 Thais bronni、麦秆虫Caprella septentrionalis |
| 生长期 | **密毛沙菜 Hypnea boergesenii、裂片石莼 Ulva fasciata | |
| 茂盛期 | **铜藻 Sargassum horneri、密毛沙菜 Hypnea boergesenii、石莼 Ulva lactuca L. |
注: **为各站点优势种(优势度IRI≥1000), 其余为各站点重要种(优势度IRI为100~1000) |
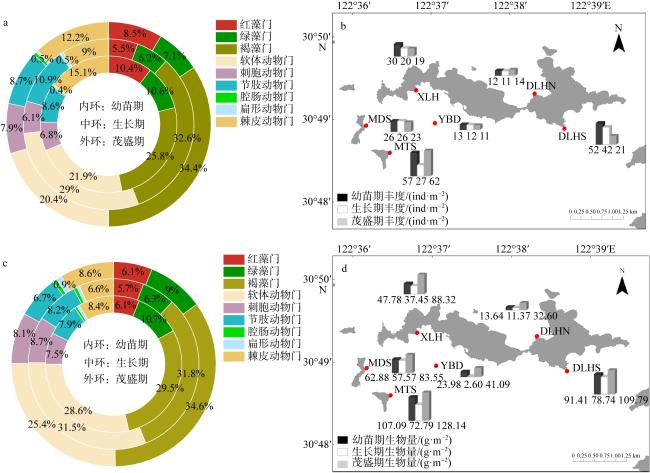
图4 不同生长时期大型底栖生物各类群平均丰度(a)和平均生物量组成(c)、各站点平均丰度(b)和平均生物量分布(d)图b、d基于自然资源部标准地图服务网站下载的审图号为GS(2024)0650号的标准地图制作 Fig. 4 Variations in macrozoobenthos groups: Average abundance (a) and average biomass composition (c) during different growth periods; average abundance (b) and average biomass (d) distribution across stations |
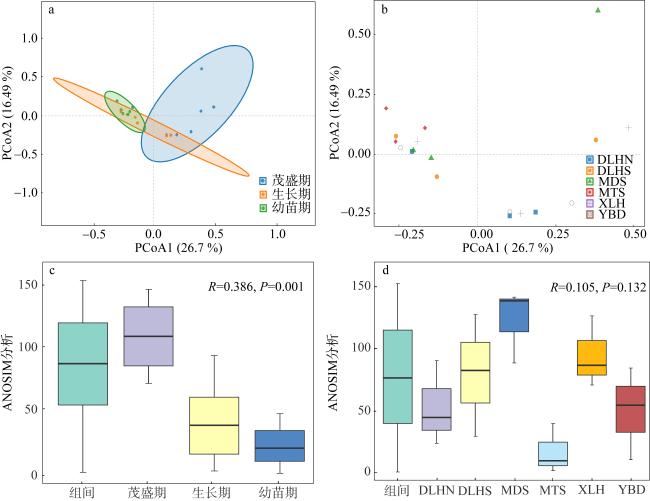
图6 大型底栖生物群落结构的PCoA与ANOSIM分析a与c为三个时期大型底栖生物群落结构差异的分析与差异检验; b与d分别为六个站点大型底栖生物群落结构差异的分析与差异检验 Fig. 6 PCoA and ANOSIM analyses of macrobenthic community structure. (a, c) Analysis and difference testing of the macrobenthic community structure differences across three periods; (b, d) analysis and difference testing of the macrobenthic community structure differences among six sites, respectively |
表2 不同站点和生长时期中4种环境因子的显著性检验Tab. 2 ANOVA significance tests of four environmental factors among different stations and growth stages |
| 站点 | 生长时期 | T/℃ | S/‰ | pH | DO/(mg·L-1) |
|---|---|---|---|---|---|
| MTS | 幼苗期 | 26.50±0.34 | 28.25±0.26 | 9.54±0.05 | 9.10±0.28 |
| 生长期 | 22.10±0.49 | 27.82±0.31 | 9.35±0.06 | 8.37±0.46 | |
| 茂盛期 | 23.51±0.15 | 28.42±0.37 | 9.27±0.06 | 7.30±0.14 | |
| DLHS | 幼苗期 | 26.91±0.18 | 28.10±0.15 | 9.57±0.04 | 7.02±0.31 |
| 生长期 | 21.97±0.13 | 27.76±0.36 | 9.33±0.06 | 8.27±0.37 | |
| 茂盛期 | 26.27±0.16 | 28.55±0.36 | 9.43±0.03 | 9.00±0.36 | |
| XLH | 幼苗期 | 26.70±0.24 | 27.21±0.19 | 9.43±0.03 | 9.00±0.11 |
| 生长期 | 21.30±0.40 | 27.65±0.36 | 9.20±0.01 | 8.37±0.32 | |
| 茂盛期 | 23.42±0.42 | 28.15±0.37 | 8.97±0.09 | 7.14±0.30 | |
| MDS | 幼苗期 | 27.40±0.18 | 28.21±0.36 | 9.46±0.04 | 9.10±0.18 |
| 生长期 | 21.50±0.35 | 27.54±0.11 | 9.31±0.06 | 8.32±0.46 | |
| 茂盛期 | 23.47±0.41 | 28.39±0.20 | 9.16±0.02 | 7.22±0.12 | |
| DLHN | 幼苗期 | 25.54±0.19 | 28.42±0.43 | 9.29±0.07 | 9.00±0.25 |
| 生长期 | 20.54±0.39 | 26.74±0.34 | 9.18±0.04 | 8.32±0.45 | |
| 茂盛期 | 25.64±0.11 | 28.38±0.15 | 9.09±0.01 | 9.22±0.15 | |
| YBD | 幼苗期 | 26.50±0.27 | 27.17±0.38 | 9.14±0.09 | 9.23±0.49 |
| 生长期 | 21.50±0.26 | 27.16±0.39 | 9.08±0.03 | 8.23±0.41 | |
| 茂盛期 | 21.96±0.16 | 27.45±0.14 | 9.06±0.04 | 7.21±0.45 | |
| ANOVA | P<0.05 | P<0.05 | P<0.05 | P=0.42 |
| [1] |
包炎琳, 段元亮, 杨娜, 等, 2020. 嵊泗列岛贻贝养殖区与无人岛潮间带大型海藻群落结构比较[J]. 海洋渔业, 42(5): 595-607.
|
| [2] |
毕远新, 章守宇, 王伟定, 等, 2014. 枸杞岛铜藻垂直分布格局及成因分析[J]. 生态学报, 34(17): 4931-4937.
|
| [3] |
柴召阳, 霍元子, 于克锋, 等, 2013. 枸杞岛瓦氏马尾藻藻场生态系统健康评价[J]. 海洋环境科学, 32(3): 386-389.
|
| [4] |
陈亮然, 章守宇, 陈彦, 等, 2015. 枸杞岛马尾藻场铜藻的生命史与形态特征[J]. 水产学报, 39(8): 1218-1229.
|
| [5] |
丁朋朋, 高春霞, 田思泉, 等, 2019. 浙江南部近海蟹类群落结构及其与环境因子的关系[J]. 海洋渔业, 41(6): 652-662.
|
| [6] |
董世淇, 张合烨, 孙国庆, 等, 2023. 北黄海褡裢岛海藻场邻近海域大型底栖动物群落营养结构的季节变化[J]. 应用生态学报, 34(7): 1763-1770.
|
| [7] |
傅海霞, 张玉洲, 黄书雅, 等, 2024. 徽水河底栖动物群落结构季节动态及构建机制[J]. 长江流域资源与环境, 33(6): 1239-1249.
|
| [8] |
胡益峰, 蒋红, 柳涛, 等, 2015. 马鞍列岛保护区海域水环境质量分区评价[J]. 海洋环境科学, 34(2): 240-244.
|
| [9] |
黄宗国, 林茂, 2012. 中国海洋生物图集. 第二册[M]. 北京: 海洋出版社: 255-305 (in Chinese).
|
| [10] |
金文育, 彭欣, 王宁, 等, 2017. 渔山列岛夏季潮间带大型底栖生物群落结构研究[J]. 海洋科学, 41(3): 17-25.
|
| [11] |
李宝泉, 李新正, 陈琳琳, 等, 2019. 中国海岸带大型底栖动物资源[M]. 北京: 科学出版社: 99-268.
|
| [12] |
李训猛, 章守宇, 王凯, 等, 2020. 周年温度变化对枸杞岛铜藻生长特性的影响[J]. 海洋与湖沼, 51(5): 1136-1143.
|
| [13] |
李志鹏, 杜震洪, 张丰, 等, 2016. 基于GIS的浙北近海海域生态系统健康评价[J]. 生态学报, 36(24): 8183-8193.
|
| [14] |
梁金玲, 汪振华, 章守宇, 等, 2016. 马鞍列岛海洋特别保护区虾类组成及其多样性[J]. 上海海洋大学学报, 25(1): 124-134.
|
| [15] |
梁静香, 周永东, 王忠明, 等, 2020. 三门湾大型底栖动物群落结构及其与环境因子的关系[J]. 应用生态学报, 31(9): 3187-3193.
|
| [16] |
廖一波, 曾江宁, 陈全震, 等, 2007. 嵊泗海岛不同底质潮间带春秋季大型底栖动物的群落格局[J]. 动物学报, 53(6): 1000-1010.
|
| [17] |
刘书荣, 周曦杰, 崔潇, 等, 2020. 贻贝筏式养殖区海藻群落对两类代表性藻栖端足目种群特征的影响[J]. 水产学报, 44(3): 461-468.
|
| [18] |
刘涛, 2017. 南海常见大型海藻图鉴[M]. 北京: 海洋出版社: 37-142 (in Chinese).
|
| [19] |
刘涛, 2018. 黄、渤海及东海常见大型海藻图鉴[M]. 北京: 海洋出版社: 43-161 (in Chinese).
|
| [20] |
刘媛媛, 张建伟, 韩军军, 等, 2015. 枸杞岛瓦氏马尾藻养殖及其对水环境因子的影响[J]. 生态学杂志, 34(11): 3214-3220.
|
| [21] |
卢占晖, 朱文斌, 徐开达, 等, 2023. 浙江沿岸海域螺贝类群落结构及其季节变化[J]. 海洋与湖沼, 54(1): 276-285.
|
| [22] |
陆艳用, 马玉心, 崔大练, 等, 2011. 中街山列岛保护区底栖海藻分布与资源特征[J]. 水产科学, 30(5): 269-275.
|
| [23] |
马文刚, 夏景全, 魏一凡, 等, 2022. 三亚蜈支洲岛海洋牧场近岛区底表大型底栖动物群落结构及评价[J]. 热带海洋学报, 41(3): 135-146.
|
| [24] |
毛翰宣, 秦诗立, 徐博文, 等, 2021. 浙江舟山群岛新区主要海岛功能布局研究[J]. 海洋开发与管理, 38(12): 46-51.
|
| [25] |
毛欣欣, 蒋霞敏, 林清菁, 2011. 浙江大型海藻彩色图集[M]. 北京: 科学出版社: 87-149.
|
| [26] |
裴文豪, 胡成业, 张秀梅, 等, 2023. 枸杞岛贻贝养殖区大型底栖动物群落结构及与环境因子的关系[J]. 海洋与湖沼, 54(2): 526-536.
|
| [27] |
彭欣, 谢起浪, 李尚鲁, 等, 2010. 浙南潮间带大型底栖藻类时空分布及多样性研究[J]. 热带海洋学报, 29(3): 135-140.
|
| [28] |
宋晨, 许丽婷, 王晓波, 等, 2022. 浙江舟山海域大型底栖动物群落组成及主要影响因子分析[J]. 海洋与湖沼, 53(6): 1585-1597.
|
| [29] |
田阔, 胡扬, 王一航, 等, 2024. 枸杞岛不同生境大型藻类与藻床无脊椎动物功能群的关联性分析[J]. 海洋与湖沼, 55(5): 1172-1182.
|
| [30] |
王航俊, 姚炜民, 林义, 等, 2020. 乐清湾大型底栖动物群落及其与环境因子之间的关系[J]. 海洋学报, 42(2): 75-86.
|
| [31] |
王蕾, 2011. 枸杞岛海藻场生态系统与鱼类群落季节变化的关系[D]. 上海: 上海海洋大学.
|
| [32] |
王蕾, 章守宇, 汪振华, 等, 2011. 枸杞岛近岸3种生境鱼类群落组成及岩礁区底栖海藻对鱼类群落结构的影响[J]. 水产学报, 35(7): 1037-1049.
|
| [33] |
王腾飞, 蒋霞敏, 王稼瑞, 等, 2013. 渔山列岛潮间带大型海藻的分布特征[J]. 海洋环境科学, 32(6): 836-840.
|
| [34] |
王铁杆, 胡仁勇, 张鹏, 等, 2012. 洞头列岛大型底栖海藻的组成与区系分布[J]. 上海海洋大学学报, 21(6): 1025-1031.
|
| [35] |
汪振华, 沈慧, 林军, 等, 2022. 马鞍列岛东部海域小黄鱼时空分布特征[J]. 水产学报, 46(6): 959-972.
|
| [36] |
吴程宏, 章守宇, 周曦杰, 等, 2017. 岛礁海藻场沉积有机物来源辨析[J]. 水产学报, 41(8): 1246-1255.
|
| [37] |
吴祖立, 章守宇, 陈彦, 等, 2015. 枸杞岛海藻场大型底栖无脊椎动物摄食类群研究[J]. 水产学报, 39(3): 381-391.
|
| [38] |
杨梅, 李新正, 徐勇, 等, 2016. 胶州湾潮下带大型底栖动物群落的季节变化[J]. 生物多样性, 24(7): 820-830.
|
| [39] |
曾呈奎, 2000. 中国海藻志: 第三卷·褐藻门·第二册·墨角藻目[M]. 北京: 科学出版社 (in Chinese).
|
| [40] |
张才学, 周伟男, 孙省利, 等, 2020. 硇洲岛大型海藻群落的季节演替[J]. 热带海洋学报, 39(1): 74-84.
|
| [41] |
张建琳, 包炎琳, 孙彬, 等, 2023. 下三横山岛潮间带大型海藻群落构成及季节变化[J]. 应用海洋学学报, 42(2): 225-234.
|
| [42] |
章守宇, 崔潇, 汪振华, 等, 2021. 枸杞岛贻贝养殖筏架附着海藻的群落结构[J]. 水产学报, 45(5): 726-739.
|
| [43] |
章守宇, 梁君, 汪振华, 等, 2008. 浙江马鞍列岛海域潮间带底栖海藻分布特征[J]. 应用生态学报, 19(10): 2299-2307.
|
| [44] |
章守宇, 刘书荣, 周曦杰, 等, 2019. 大型海藻生境的生态功能及其在海洋牧场应用中的探讨[J]. 水产学报, 43(9): 2004-2014.
|
| [45] |
章守宇, 汪振华, 林军, 等, 2007a. 枸杞岛海藻场夏、秋季的渔业资源变化[J]. 海洋水产研究, (1): 45-52.
|
| [46] |
章守宇, 孙宏超, 2007b. 海藻场生态系统及其工程学研究进展[J]. 应用生态学报, 18(7): 1647-1653.
|
| [47] |
张素萍, 张均龙, 陈志云, 等, 2016. 黄渤海软体动物图志[M]. 北京: 科学出版社: 5-384.
|
| [48] |
赵静, 梁金玲, 周曦杰, 等, 2017. 基于GAM模型的马鞍列岛海域优势甲壳类与环境因子的关系研究[J]. 南方水产科学, 13(3): 26-35.
|
| [49] |
赵玉喜, 王珍岩, 2021. 春季长江冲淡水在口门外海域逐月变化及其影响因素分析[J]. 海洋科学, 45(10): 81-92.
|
| [50] |
郑海斌, 蒋霞敏, 傅财华, 等, 2011. 浙江东极潮间带底栖海藻分布特征[J]. 宁波大学学报(理工版), 24(4): 29-35.
|
| [51] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2007. 海洋调查规范第6部分: 海洋生物调查 (GB/T 12763. 6—2007)[S]. 北京: 中国标准出版社.
General Administration of Quality Supervision, Inspection and Quarantine of the People's Republic of China, Standardization Administration of the People's Republic of China, 2007. Specification for oceanographic survey - Part 6: Marine biological survey(GB/T 12763.6—2007)[S]. Beijing: Standards press of China (in Chinese).
|
| [52] |
祝超文, 张虎, 袁健美, 等, 2022. 南黄海潮间带大型底栖动物群落组成及时空变化[J]. 上海海洋大学学报, 31(4): 950-960.
|
| [53] |
周曦杰, 2018. 藻场碎屑产生机制及岛礁生态养护功能实现途径研究[D]. 上海: 上海海洋大学.
|
| [54] |
周曦杰, 章守宇, 王旭, 等, 2015. 枸杞岛海藻场角蝾螺夏季摄食选择性及其生态学意义[J]. 水产学报, 39(4): 511-519.
|
| [55] |
邹敏, 章守宇, 周曦杰, 等, 2017. 不同溶解氧浓度下瓦氏马尾藻碎屑分解规律[J]. 上海海洋大学学报, 26(4): 554-560.
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}