
海南新村港和黎安港海草床大型底栖生物的群落结构比较*
*感谢中国科学院南海海洋研究所黄小平研究员、江志坚研究员、李开枝研究员和厦门大学蔡立哲教授团队在样品采集和鉴定工作以及论文写作中给予的帮助
|
何晨(1999−), 男, 广东省佛山市人, 硕士研究生, 研究方向: 海洋贝类分类学。email: tingsaki@outlook.com |
Copy editor: 孙翠慈
收稿日期: 2024-09-06
修回日期: 2024-10-13
网络出版日期: 2025-01-02
基金资助
国家科技基础资源调查专项(2015FY110600)
广东省科技计划项目(2023B1212060047)
广东省科技计划项目(2024B1212040008)
中国科学院分类学科学家岗位项目(CAS-TAX-24-043)
国家动物标本资源库
Comparison of macrobenthic community structure between seagrass beds in Xincungang and Li’angang, Hainan
Received date: 2024-09-06
Revised date: 2024-10-13
Online published: 2025-01-02
Supported by
National Investigation of Technological Basic Resources Project(2015FY110600)
Science and Technology Planning Project of Guangdong Province(2023B1212060047)
Science and Technology Planning Project of Guangdong Province(2024B1212040008)
Taxonomist Project, Chinese Academy of Sciences(CAS-TAX-24-043)
National Animal Collection Resource Center, China
何晨 , 王佳宁 , 周林滨 , 柯志新 , 刘炜炜 , 刘甲星 , 王军星 , 谭烨辉 , 陈志云 . 海南新村港和黎安港海草床大型底栖生物的群落结构比较*[J]. 热带海洋学报, 2025 , 44(4) : 106 -118 . DOI: 10.11978/2024170
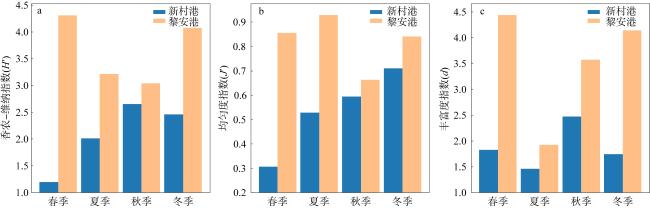
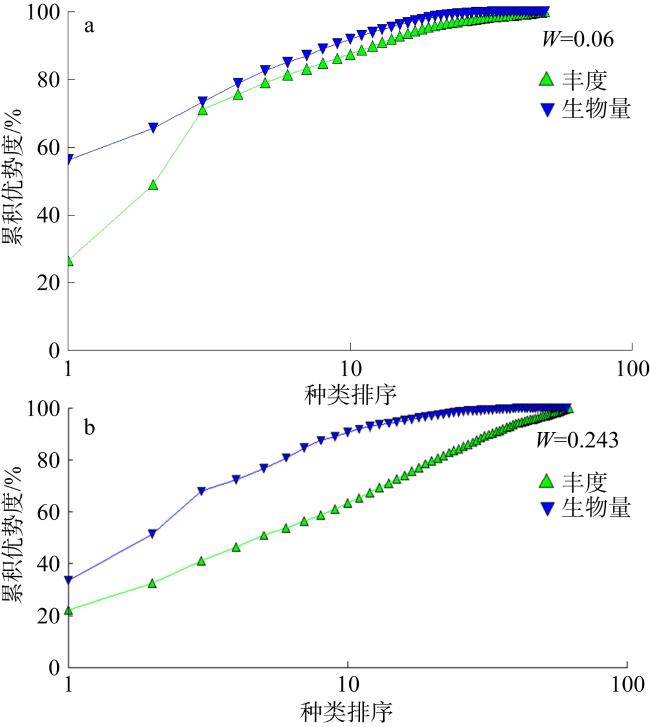
Based on the ecological survey data of macrobenthos in the seagrass beds of Xincungang (2015−2016) and Li’angang (2017−2018) across four seasons, we compared their community structures. The results showed that: (1) A total of 96 species of macrobenthos were recorded in the seagrass beds of the two areas, with 50 species in Xincungang and 62 species in Li’angang. Both seagrass beds were dominated by annelids, and Dasybranchus caducus was a common dominant species. Except in autumn, the average density of inhabited macrobenthos in Xincungang was higher than in Li’angang, while, except in summer, the biomass in Li’angang was higher than in Xincungang. (2) The Shannon-Wiener diversity index (H′), Pielou’s evenness index (J′), and Margalef’s richness index (d) in Li’angang were all higher than those in Xincungang. The W value of the abundance/biomass curves (ABC) in Li’angang was also significantly higher than in Xincungang, indicating that the macrobenthos community structure in Li’angang’s seagrass beds was more stable and less disturbed. (3) Cluster analysis of community inhabited density and biomass revealed that the communities in the two areas could be divided into two distinct groups at a similarity level of approximately 15%, demonstrating significant differences in the macrobenthos community structure between Xincungang and Li’angang. Combined with historical research, these differences are likely attributed to variations in hydrological conditions and pollution from aquaculture activities. Compared with historical data, the macrobenthos communities in the seagrass bed protected areas of both locations have shown signs of recovery, suggesting that ongoing protection and restoration efforts are effective and necessary.
Key words: seagrass beds; macrobenthos; community structure; biodiversity; Hainan
图1 新村港与黎安港采样断面图该图基于福建省自然资源厅标准地图服务系统(https://fujian.tianditu.gov.cn/map/)下载的审图号为GS(2023)336的标准地图制作, 底图无修改 Fig. 1 Location of sampling sections in Xincungang and Li’angang |
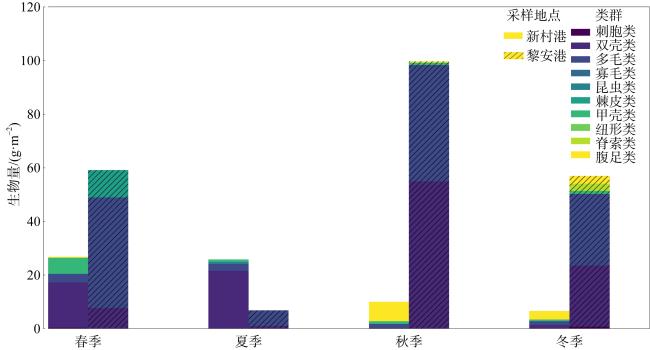
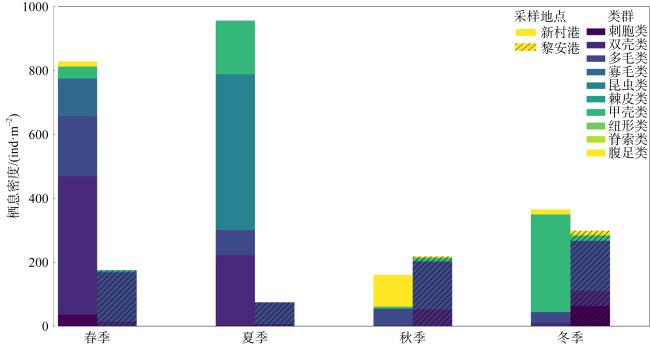
表1 新村港和黎安港各季节大型底栖生物主要类群种类数量Tab. 1 Seasonal variations in major macrobenthic groups in Xincungang and Li’angang |
| 分类阶元 | 春季种类 | 夏季种类 | 秋季种类 | 冬季种类 | ||||
|---|---|---|---|---|---|---|---|---|
| 新村港 | 黎安港 | 新村港 | 黎安港 | 新村港 | 黎安港 | 新村港 | 黎安港 | |
| 环节动物门Annelida | 15 | 18 | 8 | 6 | 5 | 13 | 9 | 15 |
| 节肢动物门Arthropoda | 4 | 0 | 2 | 0 | 2 | 2 | 2 | 3 |
| 软体动物门Mollusca | 2 | 7 | 2 | 2 | 5 | 11 | 4 | 8 |
| 刺胞动物门Cnidaria | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 0 |
| 纽形动物门Nemertea | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| 脊索动物门Chordata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| 棘皮动物门Arthropoda | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
表2 新村港和黎安港大型底栖生物不同季节的优势种及其优势度指数(Y)Tab. 2 Dominant species and their dominance indices (Y) of macrobenthos in Xincungang and Li’angang across different seasons |
| 优势种 | 春季Y | 夏季Y | 秋季Y | 冬季Y | ||||
|---|---|---|---|---|---|---|---|---|
| 新村港 | 黎安港 | 新村港 | 黎安港 | 新村港 | 黎安港 | 新村港 | 黎安港 | |
| 小头虫 Capitella capitata | 0.086 | |||||||
| 岩虫 Marphysa sanguinea | 0.025 | |||||||
| 多色天螺 Diala varia | 0.404 | |||||||
| 珠带拟蟹守螺 Cerithidea cingulata | 0.037 | |||||||
| 厚鳃蚕 Dasybranchus caducus | 0.021 | 0.092 | 0.054 | 0.119 | 0.061 | |||
| 沼蚓 Limnodriloides sp. | 0.04 | |||||||
| 南海毛满月蛤 Pillacina cietnamica | 0.45 | 0.114 | ||||||
| 蜾蠃蜚 Corophium sp. | 0.022 | |||||||
| 强壮藻钩虾 Ampithoe valida | 0.06 | 0.626 | ||||||
| 摇蚊科 Chironomidae | 0.191 | |||||||
| 软须阿曼吉虫 Arandia leptocirris | 0.042 | |||||||
| 奋镜蛤 Dosinia exasperata | 0.051 | |||||||
| 海葵 Haliplanella sp. | 0.028 | |||||||
| 背蚓虫 Notomastus sp. | 0.024 | |||||||
| 锥稚虫 Aonicles oxycephala | 0.023 | 0.03 | ||||||
| 莫桑比克岩虫 Marphysa mossabicca | 0.03 | |||||||
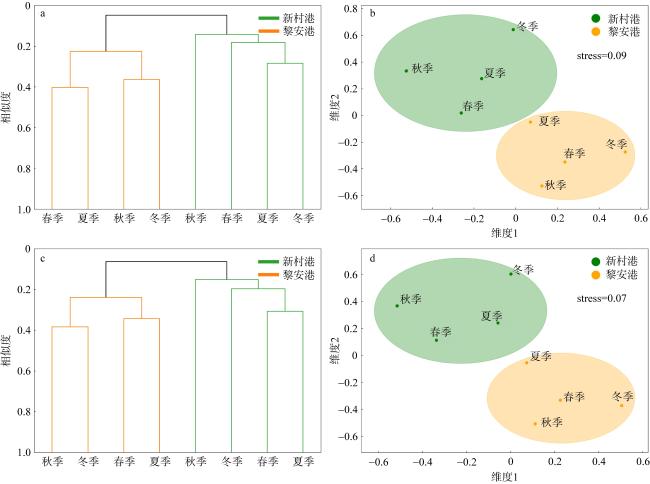
图5 基于栖息密度和生物量对新村港与黎安港大型底栖生物的UPGMA聚类和nMDS标序a. 新村港底栖生物栖息密度的UPGMA聚类; b. 新村港大型底栖生物栖息密度的nMDS标序; c. 黎安港底栖生物生物量的UPGMA聚类; d. 黎安港大型底栖生物生物量的nMDS标序 Fig. 5 UPGMA clustering and nMDS ordination based on the density and biomass of macrobenthos in Xincungang and Li’angang. (a) UPGMA clustering of macrobenthos density in Xincungang; (b) nMDS ordination of macrobenthos density in Xincungang; (c) UPGMA clustering of macrobenthos biomass in Li’angang; (d) nMDS ordination of macrobenthos biomass in Li’angang |
表3 新村港和黎安港海草床大型底栖动物主要群落参数历史数据对比Tab. 3 Comparison of key macrobenthic community parameters in seagrass beds between Xincungang and Li’angang: present vs. historical data |
| 调查结果 | 调查区域 | 物种数量 | 栖息密度均值/(ind·m-2) | 生物量均值/(g·m-2) | 多样性指数(H′) |
|---|---|---|---|---|---|
| 2002年(黄小平 等, 2007) | 新村港 | 10 | 1152.68 | 286.77 | 0.67 |
| 黎安港 | 24 | 2317.6 | 856.59 | 2.13 | |
| 2009—2013年(涂志刚 等, 2016) | 新村港和黎安港 | 95 | 137.53 | 167. 57 | 1.9 |
| 2013年(李洋, 2015) | 新村港 | 42 | 518.38 | 201.9 | 2.83 |
| 2015—2016年(本研究) | 新村港 | 50 | 576.75 | 17.27 | 2.08 |
| 2017—2018年(本研究) | 黎安港 | 62 | 191 | 222.59 | 3.66 |
| 2020年(郭治明 等, 2023) | 新村港 | 15 | 292.6 | 81.82 | 0.81 |
| 黎安港 | 17 | 133.4 | 102.22 | 0.95 |
附表1 新村港和黎安港海草床大型底栖生物名录Appendix Tab. 1 List of macrobenthos in the seagrass beds of Xincungang and Li’angang |
| 中文名 | 拉丁名 | 春季 | 夏季 | 秋季 | 冬季 | ||||
|---|---|---|---|---|---|---|---|---|---|
| 新村港 | 黎安港 | 新村港 | 黎安港 | 新村港 | 黎安港 | 新村港 | 黎安港 | ||
| 海葵目 | Actiniaria | √ | |||||||
| 柱形阿里螺 | Aliculastrum cylindricum | √ | |||||||
| 西方似蛰虫 | Amaeana occidentalis | √ | |||||||
| 似蛰虫 | Amaeana trilobata | √ | √ | ||||||
| 藻钩虾 | Ampithoe sp. | √ | |||||||
| 强壮藻钩虾 | Ampithoe valida | √ | √ | ||||||
| 无齿蛤 | Anodontia edentula | √ | √ | ||||||
| 鳞杓拿蛤 | Anomalodiscus squamosus | √ | |||||||
| 锥稚虫 | Aonides oxycephala | √ | √ | √ | |||||
| 大角玻璃钩虾 | Apohyale grandicornis | √ | |||||||
| 矮小稚齿虫 | Apoprionospio pygmaea | √ | |||||||
| 飞白枫海星 | Archaster typicus | √ | |||||||
| 凸壳肌蛤 | Arcuatula senhousia | √ | √ | √ | |||||
| 软须阿曼吉虫 | Armandia leptocirris | √ | √ | ||||||
| 中阿曼吉虫 | Armandia intermedia | √ | |||||||
| 海鞘 | Ascidia sp. | √ | |||||||
| 网纹藤壶 | Balanus inprovisus | √ | √ | ||||||
| 黑斑鳍缨虫 | Branchiomma nigromaculatum | √ | √ | ||||||
| 斑鳍缨虫 | Branchiomma sp. | √ | |||||||
| 小头虫 | Capitella capitata | √ | √ | √ | |||||
| 角沙蚕 | Ceratonereis mirabilis | √ | √ | ||||||
| 刚鳃虫 | Chaetozone setosa | √ | |||||||
| 摇蚊 | Chironomus sp. | √ | √ | ||||||
| 美女蛤 | circe scripta | √ | |||||||
| 须鳃虫 | Cirriformia tentaculata | √ | |||||||
| 奥莱彩螺 | Clithon oualaniensis | √ | |||||||
| 蜾蠃蜚 | Corophium sp. | √ | |||||||
| 厚鳃蚕 | Dasybranchus caducus | √ | √ | √ | √ | √ | |||
| 多色天螺 | Diala varia | √ | |||||||
| 奋镜蛤 | Dosinia exasperata | √ | √ | ||||||
| 矶沙蚕 | Eunice sp. | √ | |||||||
| 真裂虫 | Eusyllis sp. | √ | |||||||
| 艾裂虫亚科 | Exogoninae | √ | |||||||
| 凸加夫蛤 | Gafrarium tumidum | √ | |||||||
| 加夫蛤 | Gafrarium pectinatum | √ | √ | √ | |||||
| 钩虾 | Gammarus sp. | √ | √ | ||||||
| 皱文蛤 | Gomphina undulosa | √ | |||||||
| 海葵 | Haliplanella sp. | √ | |||||||
| 纵条矶海葵 | Haliplanella luciae | √ | √ | ||||||
| 丝异须虫 | Heteromastus sp. | √ | |||||||
| 彩虹明樱蛤 | Iridona iridescens | √ | |||||||
| 红明樱蛤 | Jitlada culter | √ | |||||||
| 后指虫 | Laonice cirrata | √ | |||||||
| 简锥虫属 | Leitoscoloplis sp. | √ | √ | ||||||
| 长锥虫属 | Leodamas sp. | √ | |||||||
| 沼蚓 | Limnodriloides sp. | √ | |||||||
| 精巧扁蛰虫 | Loimia ingens | √ | |||||||
| 索沙蚕 | Lumbrineris sp. | √ | √ | ||||||
| 似帚毛虫 | Lygdamis indicus | √ | |||||||
| 大眼蟹属 | Macrophthalmus sp. | √ | |||||||
| 印度锥稚虫 | Malacoceros indicus | √ | √ | ||||||
| 岩虫 | Marphysa sanguinea | √ | √ | ||||||
| 莫桑比克岩虫 | Marphysa mossambica | √ | √ | √ | √ | ||||
| 扁平岩虫 | Marphysa depressa | √ | √ | √ | |||||
| 尼科巴立蛤蛤 | Meropesta nicobarica | √ | |||||||
| 小健足虫 | Micropodarke dubia | √ | |||||||
| 多微稚齿虫 | Microspio multidentata | √ | |||||||
| 麦氏偏顶蛤 | Modiolus metcaifei | √ | |||||||
| 寻氏肌蛤 | Musculus senhousei | √ | |||||||
| 方斑玉螺 | Naticarius onca | √ | |||||||
| 尾刺沙蚕 | Neanthes acuminata | √ | |||||||
| 纽虫门 | Nemertea sp. | √ | |||||||
| 沙蚕属 | Nereis sp. | √ | |||||||
| 沙蚕属 | Nereis sp. | √ | |||||||
| 虹光亮樱蛤 | Nitidotellina valtonis | √ | |||||||
| 小亮樱蛤 | Nitidotellina lischkei | √ | |||||||
| 亮樱蛤 | Nitidotellina nitidula | √ | |||||||
| 背蚓虫 | Notomastus latericeus | √ | |||||||
| 背蚓虫 | Notomastus sp. | √ | |||||||
| 无背毛蛇潜虫 | Oxydromus berrisfordi | √ | |||||||
| 玛叶须虫 | Phyllodoce malmgreni | √ | |||||||
| 叶须虫 | Phyllodoce aminosa | √ | |||||||
| 珠带拟蟹守螺 | Pirenella cingulata | √ | |||||||
| 褐片阔沙蚕 | Plattynereis dumerilii | √ | |||||||
| 杜氏阔沙蚕 | Platynereis dumerilii | √ | √ | ||||||
| 蛇杂毛虫 | Poecilochaetus serpens | √ | |||||||
| 才女虫 | Polydora sp. | √ | |||||||
| 多眼虫 | Polyophthalmus cf pictus | √ | |||||||
| 梭子蟹 | Portunus sp. | √ | |||||||
| 稚齿虫 | Prionospio sp. | √ | √ | √ | √ | ||||
| 膜质伪才女虫 | Pseudopolydora kempi | √ | √ | √ | |||||
| 克氏锉棒螺 | Rhinoclavis kochi | √ | |||||||
| 南海毛满月蛤 | Rugalucina vietnamica | √ | √ | √ | √ | √ | |||
| 缨鳃虫科 | Sabellidae | √ | |||||||
| 鳞腹沟虫 | Scolelepis squamata | √ | |||||||
| 红刺尖锥虫 | Scoloplos cf rubra | √ | √ | √ | |||||
| 蟳 | Scylla sp. | √ | |||||||
| 双带蛤 | Semele sp. | √ | |||||||
| 拟箱美丽蛤 | Serratina capsoides | √ | |||||||
| 红角沙蚕 | Simplisetia erythraeensis | √ | √ | √ | |||||
| 裂虫科 | Syllidae | √ | |||||||
| 海裂虫 | Syllidia armata | √ | |||||||
| 钝缀锦蛤 | Tapes dorsatus | √ | |||||||
| 缀锦蛤 | Tapes literatus | √ | √ | ||||||
| 丽缀锦蛤 | Tapes literatus aspeersa | √ | |||||||
| 梳鳃虫 | Terebellidea stroemii | √ | |||||||
| 短桨蟹 | Thalamita sp. | √ | |||||||
| 钝齿短桨蟹 | Thalamita crenata | √ | |||||||
| 覆瓦小蛇螺蛇螺 | Thylacodes adamsii | √ | |||||||
| 鳃毛虫属 | Timarete sp. | √ | |||||||
| 火腿樱蛤 | Tonganaella perna | √ | |||||||
| 模裂虫 | Typosyllis sp. | √ | |||||||
| 欧文虫 | Owenia fusiformis | √ | |||||||
| [1] |
毕耜瑶, 许永久, 俞存根, 等, 2018. 南麂列岛海洋自然保护区岩相潮间带软体动物种类组成与数量分布[J]. 水产学报, 42(6): 902-911.
|
| [2] |
蔡立哲, 马丽, 高阳, 等, 2002. 海洋底栖动物多样性指数污染程度评价标准的分析[J]. 厦门大学学报(自然科学版), 41(5): 641-646.
|
| [3] |
蔡泽富, 陈石泉, 吴钟解, 等, 2017. 海南岛海湾与潟湖中海草的分布差异及影响分析[J]. 海洋湖沼通报, (3): 74-84.
|
| [4] |
陈娴, 李洋, 2020. 陵水黎安港水质环境分析及评价[J]. 广东化工, 47(7): 76-78, 87.
|
| [5] |
郭治明, 杨熙, 余威, 等, 2023. 华南沿海海草床分布区大型底栖动物群落特征初探[J]. 应用海洋学学报, 42(3): 469-478.
|
| [6] |
韩秋影, 施平, 2008. 海草生态学研究进展[J]. 生态学报, 28(11): 5561-5570.
|
| [7] |
胡桂坤, 秦璐璐, 李郁郁, 等, 2019. 基于ABC曲线的天津潮间带生物群落受扰动的分析[J]. 天津科技大学学报, 34(5): 57-62.
|
| [8] |
黄小平, 黄良民, 2007. 中国南海海草研究[M]. 广州: 广东经济出版社 (in Chinese).
|
| [9] |
黄小平, 江志坚, 张景平, 等, 2018. 全球海草的中文命名[J]. 海洋学报, 40(4): 127-133.
|
| [10] |
黄宗国, 林茂, 2012. 中国海洋生物图集-第四册, 2-动物界[M]. 北京: 海洋出版社 (in Chinese).
|
| [11] |
李北兴, 黎傲雪, 董建宇, 等, 2023. 湛江湾潮间带大型底栖动物的群落结构及其受干扰程度[J]. 南方水产科学, 19(2): 12-20.
|
| [12] |
李洋, 2015. 海南岛东海岸海草场大型底栖生物研究[D]. 海口: 海南大学.
|
| [13] |
林显程, 凌娟, 张燕英, 等, 2019. 海草生长的影响因素及组学技术研究进展[J]. 生物技术, 29(5): 507-511.
|
| [14] |
刘瑞玉, 2008. 中国海洋生物名录[M]. 北京: 科学出版社.
|
| [15] |
刘颖, 李进京, 陈晨, 等, 2021. 浙江象山港岛屿春、夏季潮间带大型底栖生物的群落结构特征[J]. 海洋与湖沼, 52(3): 685-696.
|
| [16] |
庞巧珠, 骆丽珍, 曾广锐, 等, 2021. 黎安港海水和沉积物中叶绿素a含量及与环境因子的关系分析[J]. 海洋湖沼通报, 43(1): 133-141.
|
| [17] |
涂志刚, 韩涛生, 陈晓慧, 等, 2016. 海南陵水新村港与黎安港海草特别保护区大型底栖动物群落结构与多样性[J]. 海洋环境科学, 35(1): 41-48.
|
| [18] |
吴钟解, 陈石泉, 王道儒, 等, 2014. 海南岛东海岸海草床生态系统健康评价[J]. 海洋科学, 38(8): 67-74.
|
| [19] |
徐凤山, 张素萍, 2008. 中国海产双壳类图志[M]. 北京: 科学出版社 (in Chinese).
|
| [20] |
杨宗岱, 1982. 中国海草的生态学研究[J]. 海洋科学, (2): 34-37.
|
| [21] |
郑凤英, 邱广龙, 范航清, 等, 2013. 中国海草的多样性、分布及保护[J]. 生物多样性, 21(5): 517-526.
|
| [22] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2007a. 海洋调查规范第6部分: 海洋生物调查 (GB/T 12763. 6—2007)[S]. 北京: 中国标准出版社.
General Administration of Quality Supervision, Inspection and Quarantine of the People's Republic of China, Standardization Administration of the People's Republic of China, 2007b. Specification for oceanographic survey-Part 6: Marine biological survey (GB/T 12763.6—2007)[S]. Beijing: Standards press of China (in Chinese).
|
| [23] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2007b. 海洋监测规范第3部分:样品采集、贮存与运输 (GB 17378. 3—2007)[S]. 北京: 中国标准出版社.
General Administration of Quality Supervision, Inspection and Quarantine of the People's Republic of China, Standardization Administration of the People's Republic of China, 2007a. The specification for marine monitoring Part 3: Sample collection, storage and transportation (GB 17378. 3—2007)[S]. Beijing: Standards press of China (in Chinese).
|
| [24] |
周立柱, 杨顶田, 尹小青, 2018. 海南新村港和黎安港非点源污染负荷估算[J]. 生态科学, 37(3): 11-20.
|
| [25] |
周毅, 江志坚, 邱广龙, 等, 2023. 中国海草资源分布现状、退化原因与保护对策[J]. 海洋与湖沼, 54(5): 1248-1257.
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}