
河口近海N2O的分布特征及微生物代谢驱动机制
|
洪义国, 教授, 从事流域近海微生物氮循环过程研究, email: yghong@gzhu.edu.cn; |
|
张宝善, 共同第一作者, 博士研究生, 从事流域近海微生物氮循环过程研究, email: 2642101135@qq.com |
Copy editor: 孙翠慈
收稿日期: 2024-12-07
修回日期: 2025-01-03
网络出版日期: 2025-01-07
基金资助
国家自然科学基金(42276130)
国家自然科学基金(42476141)
广东省粤穗联合基金重点项目(2023B1515120029)
广州市科技计划项目(2025A03J3103)
Distribution characteristics and microbial metabolic driving mechanisms of N2O in estuarine and coastal waters
Received date: 2024-12-07
Revised date: 2025-01-03
Online published: 2025-01-07
Supported by
National Natural Science Foundation of China(42276130)
National Natural Science Foundation of China(42476141)
Basic and Applied Basic Research Foundation of Guangdong Province(2023B1515120029)
Science and Technology Planning Project of Guangzhou(2025A03J3103)
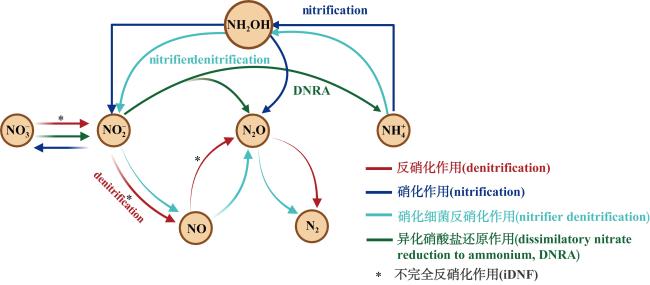
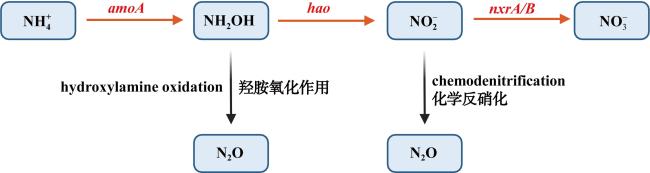
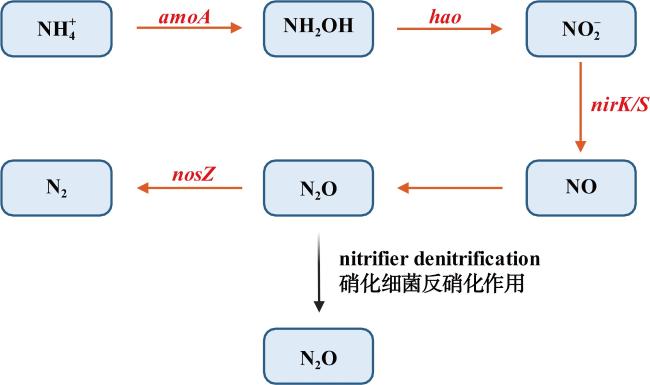
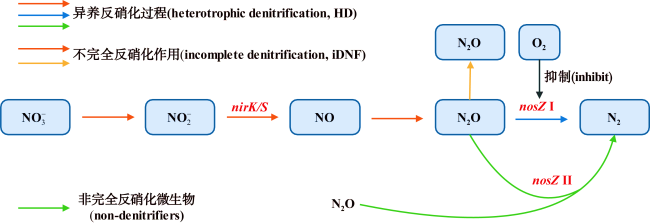
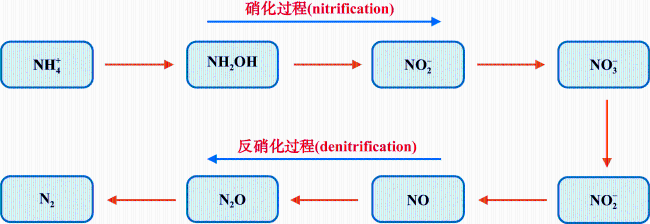
河口近海区域作为陆地与海洋生态系统的交汇点, 其在氧化亚氮(nitrous oxide, N2O)的生物地球化学循环中扮演着至关重要的角色。N2O作为一种强效温室气体, 它在河口近海的分布特征和微生物代谢驱动机制对全球气候变化具有显著影响。本文综述了河口近海区域N2O的分布特征, N2O释放的主要微生物代谢途径(硝化过程、硝化细菌反硝化过程、不完全反硝化过程、硝化作用耦合反硝化过程), N2O产生与消耗过程的测定方法以及N2O释放的环境影响因素。对河口近海N2O生物地球化学循环的研究, 有助于我们制定有效的管理策略, 对减少N2O排放、减缓全球变暖具有重要意义。
洪义国 , 张宝善 , 吴佳鹏 , 龙爱民 . 河口近海N2O的分布特征及微生物代谢驱动机制[J]. 热带海洋学报, 2025 , 44(4) : 1 -13 . DOI: 10.11978/2024229
Estuarine and coastal waters, serving as critical interfaces between terrestrial and marine ecosystems, play a pivotal role in the biogeochemical cycling of nitrous oxide (N2O), a potent greenhouse gas with significant impacts on global climate change. This review synthesizes current knowledge on the distribution patterns of N2O in estuarine and coastal regions, the key microbial metabolic pathways driving N2O emissions—including nitrification, nitrifier denitrification, incomplete denitrification, and coupled nitrification-denitrification—as well as the methodologies for quantifying N2O production and consumption. Additionally, we examine the environmental factors influencing N2O emissions. A deeper understanding of N2O biogeochemical cycling in these waters is crucial for developing effective strategies to mitigate N2O emissions and their contribution to global warming.
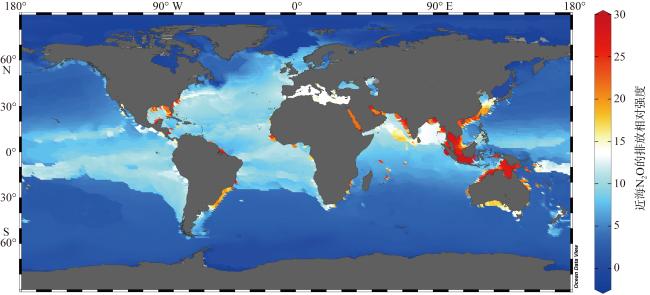
图1 近海N2O释放概念图(数据来源 https://odv.awi.de/data/ocean/world-ocean-atlas-2023/)Fig. 1 Conceptual diagram of N2O release in coastal waters (data source: https://odv.awi.de/data/ocean/world-ocean-atlas-2023/) |
| [1] |
陈玲, 白洁, 赵阳国, 等, 2016. 分离于河口区芦苇湿地1株好氧反硝化菌的鉴定及其反硝化特性[J]. 微生物学报, 56(8): 1314-1325.
|
| [2] |
丁吾举, 陆菲雨, 赵博, 等, 2024. 氮周期供给时潜流带沉积物硝化、反硝化动力学[J]. 地球科学, 49(10): 3712-3722.
|
| [3] |
洪义国, 胡耀豪, 刘晓晗, 等, 2020. 海洋N2O的排放及其关键微生物过程作用机制研究进展[J]. 微生物学报, 60(9): 1852-1864.
|
| [4] |
李冀, 朱莹, 张晓君, 2017. 非典型氧化亚氮还原酶基因nosZ Ⅱ研究进展[J]. 微生物学通报, 44(7): 1714-1719.
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
Intergovernmental Panel on Climate Change (IPCC), 2013. Climate Change 2013:The physical science basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change[R]. Cambridge and New York: Cambridge University Press.
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}