
香港牡蛎(Crassostrea hongkongensis)新型凝集素ChPerlucin的基因克隆与功能研究
|
赵泽慧(1993—), 女, 河北人, 硕士研究生, 主要从事贝类免疫与抗逆的研究。email: |
Copy editor: 姚衍桃
收稿日期: 2021-03-04
修回日期: 2021-04-19
网络出版日期: 2021-04-25
基金资助
广东省自然科学基金项目(2021A1515010541)
Molecular cloning and functional studies of ChPerlucin in Crassostrea hongkongensis
Copy editor: YAO Yantao
Received date: 2021-03-04
Revised date: 2021-04-19
Online published: 2021-04-25
Supported by
Natural Science Foundation of Guangdong Province, China(2021A1515010541)
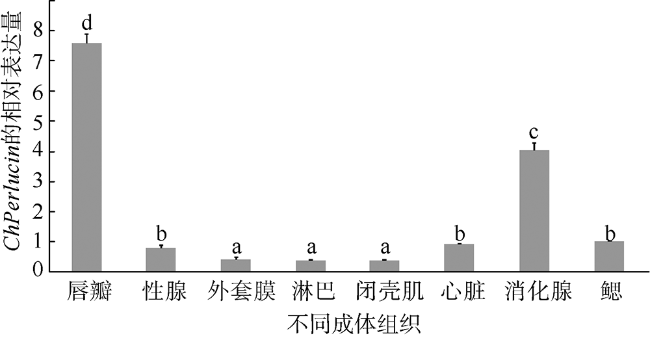
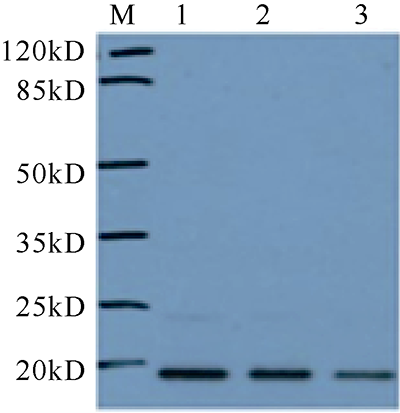
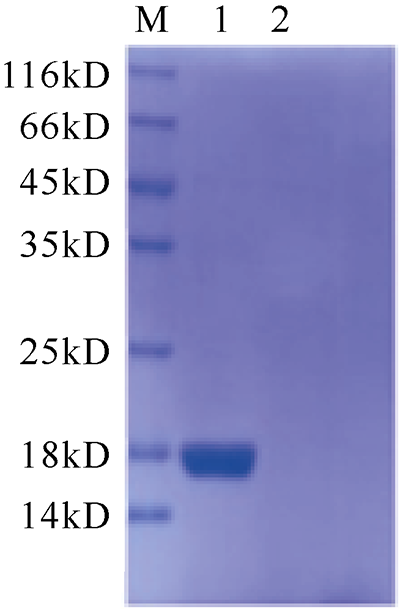
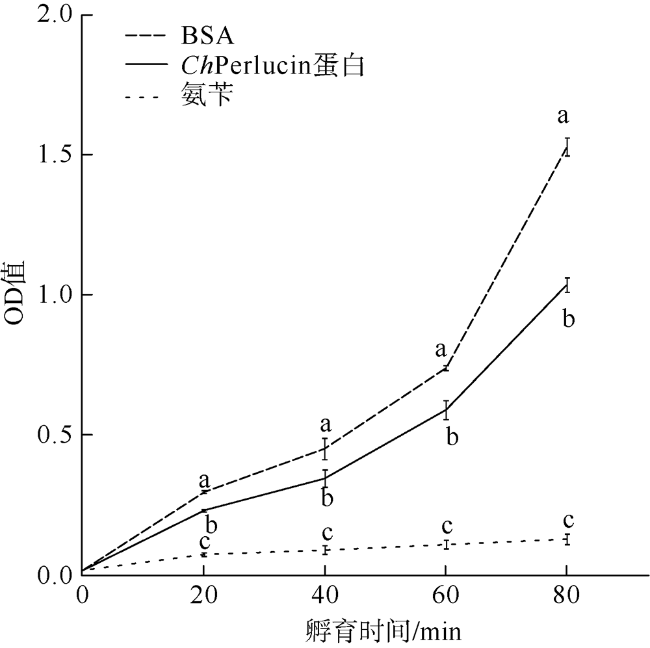
C-型凝集素(C-type lectin)是一类能识别并结合多糖的免疫蛋白质超家族, 本文首次克隆了一种新型香港牡蛎C-型凝集素基因——甘露聚糖结合凝集素, 并获得了该基因的全长序列, 命名为ChPerlucin。结果显示, ChPerlucin基因cDNA全长577bp, 包括21bp的5′末端非翻译区(untranslated region, UTR)、73bp的 3′UTR, 以及483bp的开放阅读框(open reading frame, ORF), 编码160个氨基酸, ChPerlucin蛋白理论分子量为18kD, 等电点为5.95。ChPerlucin蛋白含有保守的C-型凝集素样结构域(C-type lectin or carbohydrate-recognition domain, CLECT); 邻接法(Neighbor-Joining, NJ)进化树分析表明, ChPerlucin与其他贝类的Perlucin聚为一支, 说明该基因是软体动物Perlucin家族的新成员。qRT-PCR结果显示, ChPerlucin在香港牡蛎所检测组织和各个胚胎发育时期中均有表达; 病原菌刺激后, ChPerlucin表达量显著升高; 利用毕赤酵母真核成功表达了ChPerlucin重组蛋白, 发现ChPerlucin重组蛋白具有抑菌效果。以上结果表明ChPerlucin在香港牡蛎的先天性免疫过程中发挥了重要的作用。
关键词: 香港牡蛎; 先天性免疫; C-型凝集素; ChPerlucin
赵泽慧 , 张爱娇 , 杨雨澄 , 毛帆 , 肖述 , 李军 , 张扬 , 向志明 , 喻子牛 . 香港牡蛎(Crassostrea hongkongensis)新型凝集素ChPerlucin的基因克隆与功能研究[J]. 热带海洋学报, 2022 , 41(1) : 42 -51 . DOI: 10.11978/2021029
The C-type lectin superfamily is a class of immune proteins that can recognize and bind to polysaccharides. In this study, a new type C-type lectin gene of Crassostrea hongkongensis was cloned for the first time, and the full-length sequence of the gene was obtained. The results show that the full length of ChPerlucin gene cDNA is 577 bp, including 5′-untranslated region (UTR) in 21 bp, 3′-UTR in 73 bp, and an open reading frame (ORF) in 483 bp open reading that code 160 amino-acid polypeptide. The predicted molecular mass is 18kD, and the isoelectric point is 5.95. Amino acid sequence alignment showed that the Perlucin gene of Crassostrea hongkongensis contains a conserved carbohydrate recognition domain (CLECT). Neighbor-Joining (NJ) evolutionary tree analysis showed that ChPerlucin clustered with Perlucin from other shellfish, indicating that this gene is a new member of the Perlucin family of mollusks. The results of qRT-PCR showed that ChPerlucin was broadly expressed in various tissues and during different stages of the oyster’s embryonic and larval development; the expression of ChPerlucin increased significantly after bacteria stimulation; the recombinant protein ChPerlucin was successfully expressed using Pichia pastoris eukaryotic; and we find that ChPerlucin recombinant protein has antibacterial effect. The above results indicate that ChPerlucin plays an important role in the innate immunity of Crassostrea hongkongensis.
Key words: Crassostrea hongkongensis; innate immunity; C-type lectin; ChPerlucin
表1 引物信息Tab. 1 Sequences of designed primers used in this study |
| 引物名称 | 序列(5'-3') | 用途 |
|---|---|---|
| ChPerlucin F1 | GCTACTGCCAGGCTTACCATTCCAA | ChPerlucin 3′RACE引物 |
| ChPerlucin F2 | ATAGGAATAACGGACCTTACAGCAG | ChPerlucin 3′RACE引物 |
| ChPerlucin R1 | GTTGGATGAATACACCCACTCGTTT | ChPerlucin 5′RACE引物 |
| ChPerlucin R2 | TAAGGTCCGTTATTCCTATCCAAAA | ChPerlucin 5′RACE引物 |
| ChPerlucin F3 | GGAAAGAATGTAAGGTAAACAATGG | ChPerlucin ORF引物 |
| ChPerlucin R3 | TGACGCTTGGTTAATCAATTCTTTT | ChPerlucin ORF引物 |
| ChPerlucin F4 | CTGCCAGGCTTACCATTCCAA | ChPerlucin实时荧光定量PCR |
| ChPerlucin R4 | GGCTTGCCCAGTTTGTCGTCT | ChPerlucin实时荧光定量PCR |
| ChPerlucin F5 | AAGGATCCACCATGGGTTCCGGCTGCCGCTCGGGATGGACTCAGTACAAA | pPIC3.5K-ChPerlucin载体构建 |
| ChPerlucin R5 | TTGAATTCTCAATGATGATGATGATGATGTCCAATGATCACCGGGCCACT | pPIC3.5K-ChPerlucin载体构建 |
| GAPDH-F | GGATTGGCGTGGTGGTAGAG | GAPDH实时荧光定量PCR |
| GAPDH-R | GTATGATGCCCCTTTGTTGAGTC | GAPDH实时荧光定量PCR |
注: “F”表示正向引物, “R”表示反向引物 |
图1 ChPerlucin全长cDNA核苷酸序列及其编码的氨基酸序列(a)和CLECT结构域(b)小写字母表示UTR区, 大写字母表示ORF区, 起始密码子ATG和终止密码子TAG为加粗字体, 结构域CLECT用方框标出。*代表蛋白翻译终止, 最后连续的小写字母a表示polyA尾 Fig. 1 Nucleotide and amino acid sequences of ChPerlucin (a) and CLECT domain (b). Lower-case letters indicate UTR area, and upper-case letters indicate ORF area. Start codon ATG and stop codon TAG are bold, and the structure domain CLECT has been marked with a box. * represents the termination of protein translation, and the last continuous lower-case letter a represents the polyadenylation tail |
图2 香港牡蛎与其他物种ChPerlucin氨基酸序列的多重比对Fig. 2 Multiple alignment of the putative amino acid sequences of ChPerlucin from Crassostrea hongkongensis with other organisms homologues. The conservative cysteine sites are marked with a black five-pointed star, and the conservative EPN and WVD sites are marked with a red box. The shaded part of the same color indicates the same part, and "---" represents the amino acid deletion. Gene Bank serial number (NCBI): Crassostrea hongkongensis(MW666179); Crassostrea gigas(XP_ 011424364.2); Crassostrea virginica(XP_022310854.1); Pecten maximus(XP_033732131.1); Mizuhopecten yessoensis(XP_ 021347615.1); Argopecten irradians(ACS72239.1); Haliotis laevigata(CBK19533.1); Haliotis discus discus(ABO26595.1); Haliotis diversicolor(AEQ16381.1); Ruditapes philippinarum(ACU83224.1); Aplysia californica(XP_012943835.2); Apostichopus japonicus(BBA19537.1); Drosophila elegans(XP_017112449.1); Homo sapiens (sp|Q8IZS7.1|) |
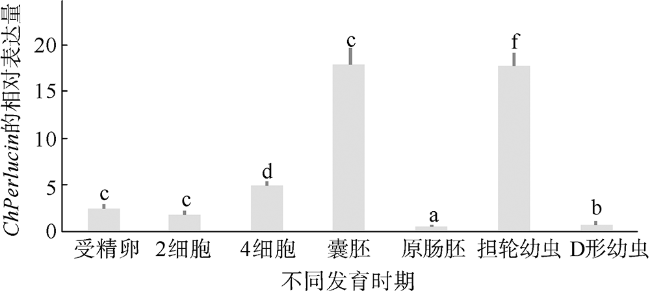
图4 ChPerlucin在不同胚胎发育时期的表达模式表达量数据为每个样品3个重复的平均值±标准差, 不同的小写字母表示差异显著(P<0.05) Fig. 4 Expression pattern of ChPerlucin in different embryonic development stages. We show the mean ± standard deviation of three replicates, and use different lower-case letters to indicate significant differences (P<0.05) |
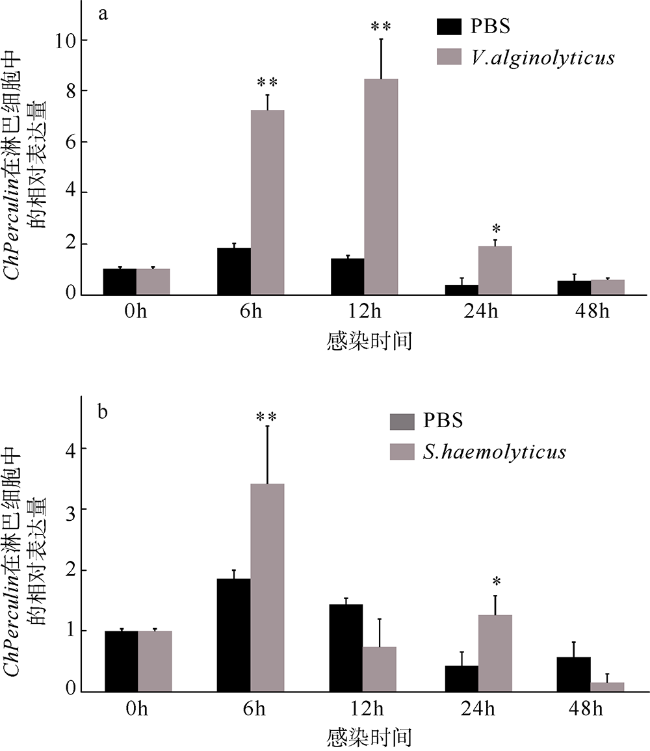
图6 ChPerlucin在细菌感染下的表达模式表达量数据为每个样品3个重复的平均值±标准差, 菌刺激下不同时间点的表达量均以对照(0h)的倍数表示, *代表与对照差异显著(P<0.05), **代表与对照差异极显著(P<0.01) Fig. 6 Expression profiles of ChPerlucin in hemocytes tissue under bacterial stress. We show the average ± standard deviation of three replicates. The expression levels at different times under bacterial stimulation are expressed as multiples of the control (0h), * represents a significant difference from the control (P<0.05), and ** represents the control difference is extremely significant (P<0.01) |
| [1] |
方紫妍, 2016. 中华绒螯蟹C型凝集素(EsLecB)的抗菌功能研究[D]. 上海: 华东师范大学.
|
| [2] |
顾圆圆, 夏正坤, 2008. 甘露糖结合凝集素的研究进展[J]. 医学研究生学报, 21(5): 537-541.
|
| [3] |
刘逸尘, 刘丽静, 张亦陈, 等, 2012. 中国明对虾C-型凝集素基因(Fclectin)的重组表达及活性分析[J]. 水产学报, 36(10): 1493-1502.
|
| [4] |
农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会, 2019. 中国渔业统计年鉴2019[M]. 北京: 中国农业出版社: 139.
Fishery Administration of the Ministry of Agriculture and Rural Areas, National Fisheries Technology Extension Center, China Society of Fisheries, 2019. China fishery statisical yearbook[M]. Beijing: China Agriculture Press: 139. (in Chinese)
|
| [5] |
孙盛明, 傅洪拓, 宣富君, 等, 2019. 日本沼虾C型凝集素结构域家族3的cDNA克隆、原核表达和定位分析[J]. 水产学报, 43(11): 2317-2326.
|
| [6] |
王昊, 2006. 栉孔扇贝凝集素家族基因的研究[D]. 青岛: 中国科学院大学(中国科学院海洋研究所).
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
/
| 〈 |
|
〉 |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}