
近40年永兴岛造礁石珊瑚动态变化及其控制因素
|
吴绵璇(1999—), 女, 广东省汕头市人, 主要从事珊瑚礁生态研究。email: 502022270103@smail.nju.edu.cn |
Copy editor: 林强
收稿日期: 2025-02-06
修回日期: 2025-03-21
网络出版日期: 2025-03-31
基金资助
中国科学院学部咨询评议项目(2016ZWH005A-005)
中国科学院学部咨询评议项目(2018-G01-B-005)
中国南海研究协同创新中心项目
江苏省自然资源发展专项资金(海洋科技创新)项目(JSZRHYKJ202104)
Dynamics and controlling factors of scleractinian corals in Yongxing Island over the past 40 years
Received date: 2025-02-06
Revised date: 2025-03-21
Online published: 2025-03-31
Supported by
Consultation and Evaluation Projects of the Chinese Academy of Sciences(2016ZWH005A-005)
Consultation and Evaluation Projects of the Chinese Academy of Sciences(2018-G01-B-005)
Project of the Collaborative Innovation Center of South China Sea Studies
Special Fund for Natural Resources Development of Jiangsu Province (Marine Science and Technology Innovation) Project(JSZRHYKJ202104)
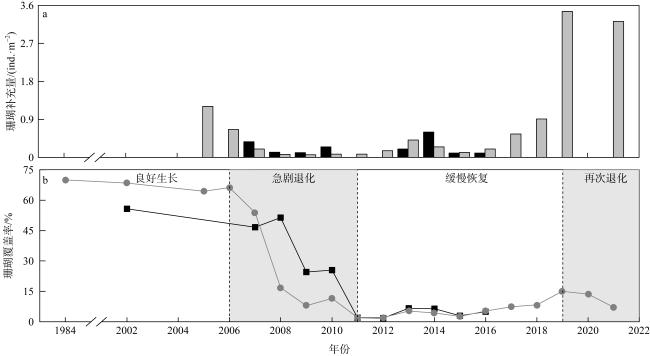
全球珊瑚礁正持续退化, 通过有效降低区域性珊瑚礁退化造成的地区压力, 以减轻日益增加的全球压力的影响, 现已成为共识。南海西沙群岛珊瑚礁是“印度洋—太平洋珊瑚礁区”的重要组成部分, 在近年来的珊瑚礁生态监测调查中, 呈现出不同程度的退化现象。准确把握其动态变化与主控因素, 是对珊瑚礁区进行及时有效管理的重要基础。本文梳理了1984—2021年西沙永兴岛珊瑚礁调查获取的珊瑚覆盖率、补充量及白化率等生态指标数据, 统计分析表明: 近40年来, 西沙永兴岛造礁石珊瑚总体呈现“良好生长—急剧退化—缓慢恢复—再次退化”的4阶段动态变化, 1984—2006年呈良好生长状态, 2006—2011年出现急剧退化阶段, 2011—2019年缓慢恢复, 2019—2021年又重新退化。长棘海星和珊瑚热白化分别是永兴岛造礁石珊瑚发生急剧退化和再次退化的主要控制因素, 珊瑚疾病在急剧退化阶段产生了一定的影响, 人类活动一定程度上抑制了珊瑚急剧退化后的迅速恢复, 同期珊瑚热白化影响了其后续恢复。相比大堡礁造礁石珊瑚急剧退化后持续恢复的变化状况, 永兴岛造礁石珊瑚退化程度更为严重, 后续10年都呈现低速恢复状态。
吴绵璇 , 罗孝文 , 张永战 . 近40年永兴岛造礁石珊瑚动态变化及其控制因素[J]. 热带海洋学报, 2025 , 44(6) : 51 -63 . DOI: 10.11978/2025019
Global coral reefs are undergoing continuous degradation, and it is universally acknowledged that alleviating local pressures stemming from regional coral reef degradation is crucial to counteracting the substantial impacts of escalating global stressors. The coral reefs of the Xisha Islands in the South China Sea, a vital component of the “Coral Triangle”, have exhibited varying degrees of degradation in recent years, as revealed by ecological monitoring. Analyzing coral reef degradation through relevant ecological indicators is vital for understanding coral reef trends and supporting restoration and management efforts. This paper, drawing upon existing surveys of Yongxing Island in the Xisha Islands, examines relevant ecological indicators such as coral coverage, replenishment, and bleaching rates. The results indicate that over the past 40 years, the scleractinian corals of Yongxing Island have undergone a phased pattern of degradation: healthy growth (1984-2006), rapid degradation (2006-2011), slow recovery (2011-2019), and renewed degradation (2019-2021). This trend follows a general pattern of “healthy growth→sharp degradation→slow recovery→renewed degradation”. The crown-of-thorns starfish emerges as the primary driver of rapid degradation, with coral diseases also playing a role during this stage. Human activities hindered the swift recovery of corals following the initial rapid degradation to a certain extent, while coral bleaching was the primary factor governing both slow recovery and later-stage renewed degradation. In comparison to the swift recovery of scleractinian corals in the Great Barrier Reef following its rapid degradation, the scleractinian corals of Yongxing Island have experienced more severe degradation, with a lower recovery rate over the past decade.
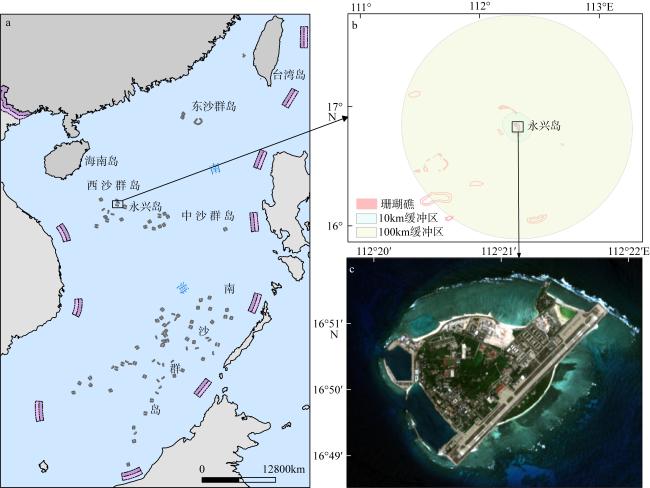
图1 永兴岛区域示意图a. 永兴岛的位置示意图; b. 永兴岛的10km和100km缓冲区范围图; c. 永兴岛的区域示意图。a基于2018年自然资源部标准地图服务网站下载的审图号为GS(2022)4318 号的标准地图制作, 底图无修改; b、c基于联合国环境规划署世界保护监测中心(UNEP-WCMC)全球珊瑚礁数据库(Version 4.1, 2021)和2024年哨兵2号(Sentinel-2)影像制作 Fig. 1 Schematic diagram of Yongxing Island in the South China Sea. (a) Location map of Yongxing Island; (b) 10 km and 100 km buffer zones of Yongxing Island; (c) regional diagram of Yongxing Island |
表1 数据来源表Tab. 1 Data sources |
表2 永兴岛及东南侧造礁石珊瑚覆盖率和补充量年变化Tab. 2 Annual variations in scleractinian coral coverage and recruitment at Yongxing Island and its southeastern side |
| 年份 | 东南侧覆盖率/% | 东南侧补充量/(ind.·m-2) | 永兴岛覆盖率/% | 永兴岛补充量/(ind.·m-2) |
|---|---|---|---|---|
| 1984 | —— | —— | 70.00 | —— |
| 2002 | 55.7 | —— | 68.40 | —— |
| 2005 | —— | —— | 64.39 | 1.21 |
| 2006 | —— | —— | 66.25 | 0.66 |
| 2007 | 46.67 | 0.37 | 53.80 | 0.20 |
| 2008 | 51.40 | 0.12 | 16.84 | 0.07 |
| 2009 | 24.50 | 0.11 | 7.93 | 0.06 |
| 2010 | 25.50 | 0.25 | 11.55 | 0.07 |
| 2011 | 1.97 | 0.00 | 2.15 | 0.07 |
| 2012 | 1.79 | 0.00 | 2.15 | 0.16 |
| 2013 | 6.67 | 0.20 | 5.25 | 0.41 |
| 2014 | 6.40 | 0.60 | 4.38 | 0.25 |
| 2015 | 3.00 | 0.10 | 2.65 | 0.12 |
| 2016 | 5.00 | 0.10 | 5.25 | 0.20 |
| 2017 | —— | —— | 7.45 | 0.55 |
| 2018 | —— | —— | 8.32 | 0.91 |
| 2019 | —— | —— | 14.99 | 3.46 |
| 2020 | —— | —— | 13.60 | —— |
| 2021 | —— | —— | 7.22 | 3.22 |
图2 1984—2021年永兴岛及东南侧造礁石珊瑚覆盖率和补充量年际变化a. 永兴岛及东南侧造礁石珊瑚补充量年变化; b. 永兴岛及东南侧造礁石珊瑚覆盖率年变化 Fig. 2 Interannual variations in scleractinian coral coverage and recruitment at Yongxing Island and its southeastern side from 1984 to 2021. (a) Annual variations in scleractinian coral recruitment; (b) annual variations in scleractinian coral coverage |
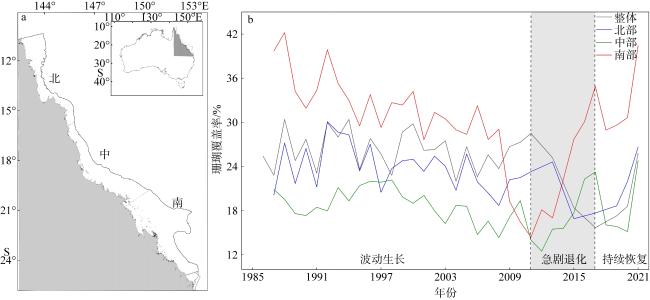
图3 1986—2021年澳大利亚大堡礁珊瑚覆盖率年际变化a. 澳大利亚大堡礁的研究区示意图; b. 澳大利亚大堡礁整体及北、中、南部珊瑚覆盖率变化 Fig. 3 Interannual variations in coral coverage at Australia’s Great Barrier Reef from 1986 to 2021. (a) Schematic map of the study area; (b) variations in coral coverage for the entire reef and its northern, central and southern sections |
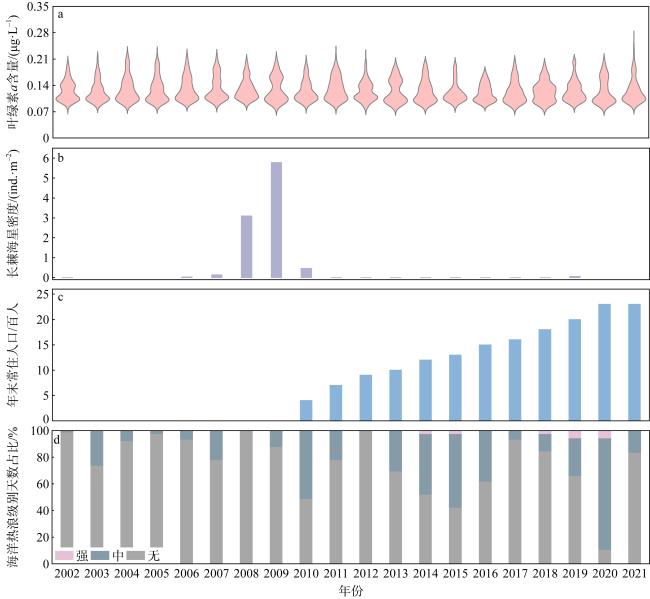
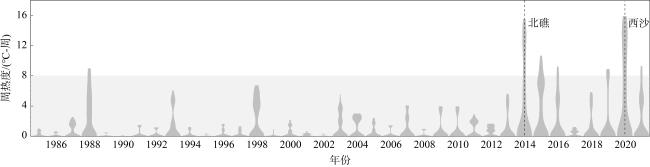
图4 2002—2021年永兴岛各退化控制因素指标年际变化a. 永兴岛叶绿素a含量; b. 长棘海星密度; c. 年末常住人口数; d. 海洋热浪级别天数占比 Fig. 4 Interannual variations in degradation control factors at Yongxing Island from 2002 to 2021. (a) Chlorophyll-a concentration; (b) density of crown-of-thorns starfish; (c) year-end resident population; (d) percentage of days with marine heatwave levels |
| [1] |
海南省海洋与渔业厅, 2013. 2012年海南省海洋环境状况公报[N]. 海南日报, 06-27(A07) (in Chinese).
|
| [2] |
黄晖, 练健生, 黄小平, 等, 2006. 用珊瑚覆盖率作为干扰指标: 永兴岛石珊瑚生物多样性研究[J]. 科学通报, 51(S3): 108-113 (in Chinese).
|
| [3] |
黄晖, 尤丰, 练健生, 等, 2011. 西沙群岛海域造礁石珊瑚物种多样性与分布特点[J]. 生物多样性, 19(6): 710-715.
|
| [4] |
黄晖, 2021. 中国珊瑚礁状况报告(2010—2019)[M]. 北京: 海洋出版社.
|
| [5] |
黄晖, 俞晓磊, 黄林韬, 等, 2024. 珊瑚礁生态学研究现状和展望[J]. 热带海洋学报, 43(3): 3-12.
|
| [6] |
旷芳芳, 张俊鹏, 查晶, 等, 2023. 不同SST资料在海南岛和北部湾珊瑚白化预警中的适用性比较[J]. 应用海洋学学报, 42(3): 371-381.
|
| [7] |
郎尚昆, 2022. 海南岛和西沙群岛珊瑚白化状况及原因分析[D]. 三亚: 海南热带海洋学院.
|
| [8] |
李淑, 余克服, 2007. 珊瑚礁白化研究进展[J]. 生态学报, 27(5): 2059-2069.
|
| [9] |
李颖虹, 黄小平, 岳维忠, 2004. 西沙永兴岛环境质量状况及管理对策[J]. 海洋环境科学, 23(1): 50-53.
|
| [10] |
李元超, 黄晖, 董志军, 等, 2008. 珊瑚礁生态修复研究进展[J]. 生态学报, 28(10): 5047-5054.
|
| [11] |
李元超, 韩有定, 陈石泉, 等, 2015. 砗磲采挖对珊瑚礁生态系统的破坏: 以西沙北礁为例[J]. 应用海洋学学报, 34(4): 518-524.
|
| [12] |
李元超, 吴钟解, 陈石泉, 等, 2017. 永兴岛及七连屿浅水礁区珊瑚礁鱼类多样性探讨[J]. 海洋环境科学, 36(4): 509-516.
|
| [13] |
李元超, 陈石泉, 郑新庆, 等, 2018. 永兴岛及七连屿造礁石珊瑚近10年变化分析[J]. 海洋学报, 40(8): 97-109.
|
| [14] |
李元超, 吴钟解, 梁计林, 等, 2019a. 近15年西沙群岛长棘海星暴发周期及暴发原因分析[J]. 科学通报, 64(33): 3478-3484.
|
| [15] |
李元超, 梁计林, 吴钟解, 等, 2019b. 长棘海星的暴发及其防治[J]. 海洋开发与管理, 36(8): 9-12.
|
| [16] |
龙笛, 2024. OCNET 全球每日叶绿素-a产品数据集(2001-2021年)[Z/OL]. (2024-07-27)[2025-0205]. 国家冰川冻土沙漠科学数据中心. https://cstr.cn/CSTR:11738.11.NCDC.ZENODO.DB6671.2024
|
| [17] |
陆永强, 陈正华, 余克服, 等, 2022. 1985-2019年南海诸岛珊瑚礁区热压力时空变化研究分析[J]. 海洋学报, 44(11): 179-190.
|
| [18] |
时小军, 刘元兵, 陈特固, 等, 2008. 全球气候变暖对西沙、南沙海域珊瑚生长的潜在威胁[J]. 热带地理, 28(4): 342-345+368.
|
| [19] |
王丽荣, 余克服, 赵焕庭, 等, 2014. 南海珊瑚礁经济价值评估[J]. 热带地理, 34(1): 44-49.
|
| [20] |
王雪辉, 杜飞雁, 林昭进, 等, 2011. 西沙群岛主要岛礁鱼类物种多样性及其群落格局[J]. 生物多样性, 19(4): 463-469+501-504.
|
| [21] |
吴程宏, 刘维, 符丹凤, 等, 2023. 西沙群岛海域砗磲资源的调查[J]. 热带生物学报, 14(2): 197-202.
|
| [22] |
吴钟解, 王道儒, 涂志刚, 等, 2011. 西沙生态监控区造礁石珊瑚退化原因分析[J]. 海洋学报, 33(4): 140-146.
|
| [23] |
许强, 刘维, 高菲, 等, 2018. 发展中国南海热带岛礁海洋牧场——机遇、现状与展望[J]. 渔业科学进展, 39(5): 173-180.
|
| [24] |
姚秋翠, 余克服, 廖芝衡, 等, 2022. 棘冠海星暴发及其对珊瑚礁的生态影响研究进展[J]. 生态学报, 42(18): 7517-7528.
|
| [25] |
余克服, 宋朝景, 赵焕庭, 1995. 西沙群岛永兴岛地貌与现代沉积特征[J]. 热带海洋, 14(2): 24-31.
|
| [26] |
袁钰, 徐海明, 马静, 等, 2024. AMO对ENSO与初夏西太平洋海洋热浪年际关系的年代际调制作用[J]. 热带海洋学报, 43(5): 1-16.
|
| [27] |
曾昭璇, 梁景芬, 丘世钧, 1997. 中国珊瑚礁地貌研究[M]. 广州: 广东人民出版社:1-474.
|
| [28] |
赵焕庭, 王丽荣, 2017. 南海诸岛珊瑚礁人工岛建造研究[J]. 热带地理, 37(5): 681-693.
|
| [29] |
朱志雄, 周永灿, 柯韶文, 等, 2012. 西沙群岛造礁石珊瑚主要疾病调查与初步研究[J]. 海洋学报, 34(6): 195-204.
|
| [30] |
邹仁林, 2001. 中国动物志:腔肠动物门, 珊瑚虫纲, 石珊瑚目, 造礁石珊瑚[M]. 北京: 科学出版社: 1-289.
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}