
海洋弧菌HN897 β-琼脂糖酶基因vas1-1339异源表达及活性分析
|
喻飞(1990— ), 男, 讲师, 博士, 研究方向为海洋微生物。email: |
收稿日期: 2021-05-21
修回日期: 2021-06-24
网络出版日期: 2021-06-30
基金资助
国家自然科学基金(31872597)
Heterologous expression and enzymatic characterization of marine Vibrio astriarenae-derived β-Agarase gene vas1-1339
Received date: 2021-05-21
Revised date: 2021-06-24
Online published: 2021-06-30
Supported by
National Natural Science Foundation of China(31872597)
Jiangsu Agriculture Science and Technology Innovation Fund(CX(19)2033)
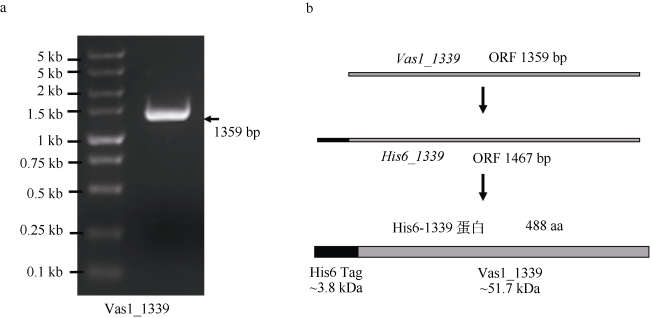
海洋弧菌HN897株是一株高产琼脂糖酶的Vibrio astriarenae菌, 前期功能缺失实验证明其β-琼脂糖酶基因vas1-1339的缺失可显著降低弧菌HN897株的琼脂水解活性。文章在大肠杆菌中异源表达Vas1-1339基因编码蛋白, 分析该基因的表达及蛋白功能活性。首先, 构建含vas1-1339基因开放阅读框全长的表达载体pET28a-1339, 利用原核表达技术在大肠杆菌中成功地异源表达了Vas1-1339琼脂糖酶; 然后, 利用卢戈氏碘液染色分析发现异源表达Vas1-1339琼脂糖酶的大肠杆菌能够高效水解琼脂, 且异丙基硫代半乳糖苷(IPTG)最优诱导浓度为10μmol·L-1; 免疫印迹分析发现胞内基因表达产物羧基端可能存在切割现象, 推测Vas1-1339琼脂糖酶存在复杂的成熟过程。对纯化的琼脂糖酶活性分析发现海洋弧菌HN897株β-琼脂糖酶Vas1-1339能够独立发挥琼脂糖降解功能。
喻飞 , 金兴坤 , 雷天影 , 曹海航 , 陈锵辉 , 阳耀帆 , 李佳航 , 赵哲 . 海洋弧菌HN897 β-琼脂糖酶基因vas1-1339异源表达及活性分析[J]. 热带海洋学报, 2022 , 41(2) : 170 -176 . DOI: 10.11978/2021065
The marine Vibrio HN897 strain is an agarose-producing Vibrio astriarenae. Studies indicated that the absence of the gene Vas1-1339, encoding a β-agarase, significantly reduced the hydrolysis effect of Vibrio HN897 strain on agarose. Herein, we further analyzed the expression and enzymatic characterization of the β-agarase gene in heterologous bacteria, Escherichia coli (E. coli). Vas1-1339 agarase was successfully expressed in E. coli, and the optimal concentration of inducer, IPTG, was at 10 μmol·L-1. Immunoblotting analysis showed that the C-terminal of the gene expression product might be cleavage in E. coli, suggesting complex maturation process of this protein. Lugol’s iodine staining analysis revealed that E. coli, expressing Vas1-1339 agarase, was highly effective in degrading agarose, and purified protein also had this similar function, which suggests that the β-agarase Vas1-1339 of the HN897 strain can independently play the role on agarolytic degradation. These results lay a preliminary foundation for the functional study and related technology application of β-agarase derived from marine vibrion.
Key words: β-agarase; marine Vibrio; agarose hydrolysis; protein expression
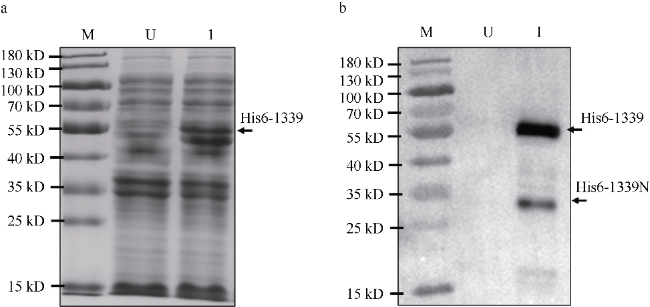
图2 重组β-琼脂糖酶His6-1339诱导表达a. β-琼脂糖酶His6-1339诱导表达样品的SDS-PAGE分析, U未诱导, I诱导; b. 图a中样品进行免疫印迹分析, 鼠抗His-tag单克隆抗体用于识别His6标签蛋白 Fig. 2 β-Agarase His6-1339 induced to express in Escherichia coli. (a) His6-tagged protein was induced from E. coli. Representative images from an SDS-PAGE analysis of His6-1339 is shown. Lanes: (U) non-induced; and (I) induced. (b) A western blot analysis of the His6-tag protein (in a) probed with a monoclonal antibody against His6-tag |
图3 卢戈氏碘液染色分析His6-1339菌株水解琼脂能力不同菌株(His6-1339和His6)培养12h后进行卢戈氏碘液染色分析, 诱导表达中IPTG使用浓度分别为0.1、1、10和100 μmol·L-1, 未加IPTG作为对照组(0μmol·L-1) Fig. 3 Lugol’s iodine staining analysis indicating activity of β-Agarase in Escherichia coli. Agar plate with E. coli strains (His6-1339 and His6) was stained by Lugol’s iodine. IPTG (0.1, 1, 10 and 100 μmol·L-1) were used to induce protein expression, and 0 μmol·L-1 IPTG served as a negative control |
图4 重组β-琼脂糖酶His6-1339诱导表达时间优化不同时间点(0、1、2、3、4 h)收集诱导菌株(His6-1339)样品进行免疫印迹分析, 鼠抗His-tag单克隆抗体用于识别His6标签蛋白, 样品IPTG使用浓度为10μmol·L-1 Fig. 4 Optimization of β-Agarase His6-1339 expression induced by IPTG. A western blot analysis of the His6-1339 protein probed with a monoclonal antibody against His-tag. The samples were collected at the indicated times, and the 10 μmol·L-1 IPTG was used to induce protein expression |
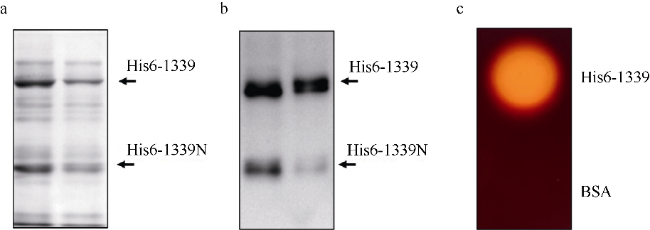
图5 纯化β-琼脂糖酶His6-1339降解琼脂糖分析a. 纯化His6-1339蛋白进行SDS-PAGE分析, 两泳道代表不同批次纯化的蛋白样品; b. 纯化的His6-1339蛋白样品进行免疫印迹分析, 鼠抗His-tag单克隆抗体用于识别His6标签蛋白, 两泳道表示不同批次纯化的蛋白样品; c. 卢戈氏碘液染色分析纯化的His6-1339蛋白水解琼脂能力, His6-1339蛋白使用量为500ng, 等量牛血清蛋白(BSA)作为对照 Fig. 5 Agarose hydrolysis of purified His6-1339 protein. (a) Purified His6-1339 protein analyzed by SDS-PAGE assay; (b) A western blot analysis of purified His6-1339 protein probed with a monoclonal antibody against His6-tag; (c) Agar plates with purified His6-1339 protein (500 ng) were stained by Lugol’s iodine. The 500 ng BSA protein serves as a control |
| [1] |
林福娣, 2020. 新琼寡糖的酶法制备及其生物活性研究[D]. 厦门: 华侨大学, 14-29.
|
| [2] |
张静雅, 刘宇鹏, 吴超, 等, 2018. 1株高产琼脂糖酶海洋弧菌的分离与酶活性的测定[J]. 热带生物学报, 9(2): 142-146.
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}