
丛生盔形珊瑚光合生理及共生真菌群落分析
|
朱文涛(1995—), 男, 安徽省安庆市人, 博士研究生。email: |
收稿日期: 2021-05-19
修回日期: 2021-07-16
网络出版日期: 2021-07-26
基金资助
国家自然科学基金(42076108)
国家自然科学基金(42161144006)
海南省重点研发计划(ZDYF2020177)
Analysis of photosynthetic physiology and symbiotic fungi community in Galaxea fascicularis
Received date: 2021-05-19
Revised date: 2021-07-16
Online published: 2021-07-26
Supported by
National Natural Science Foundation of China(42076108)
National Natural Science Foundation of China(42161144006)
Hainan Provincial Key Research and Development Program(ZDYF2020177)
Foundation of Hainan University(KYQD(ZR)1805)
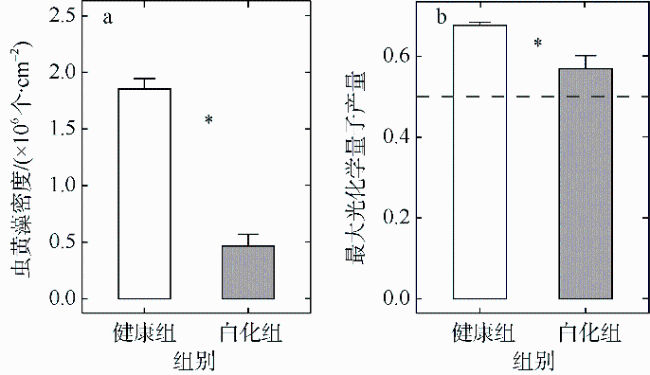
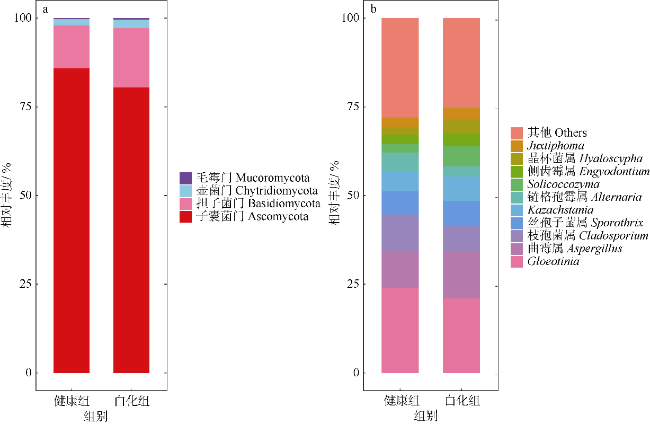
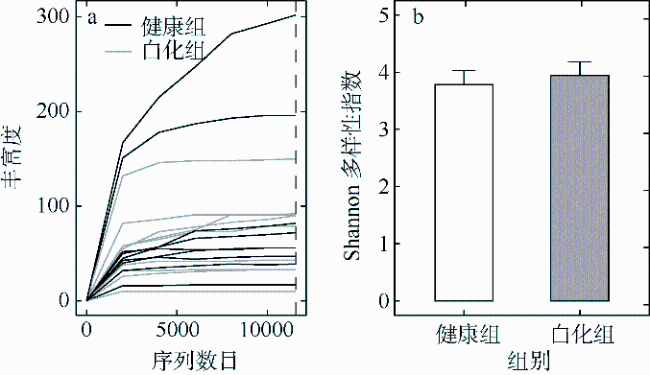
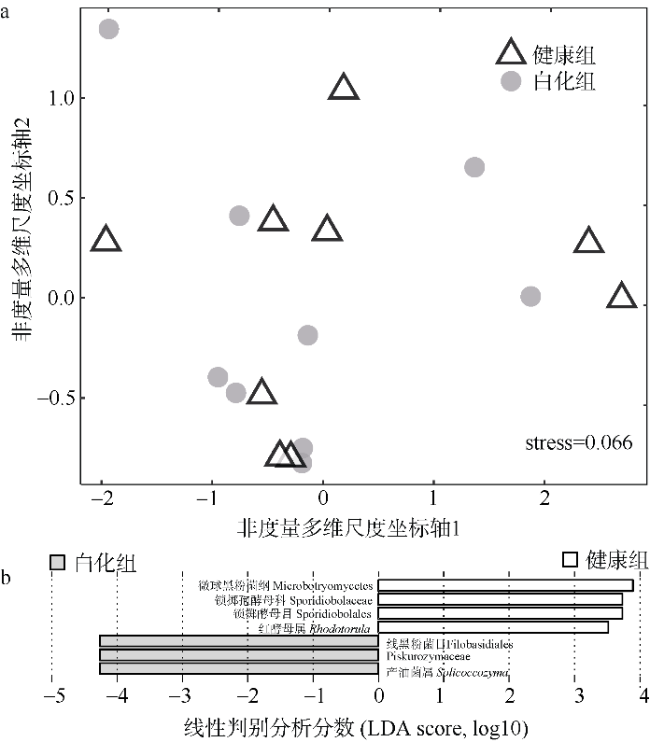
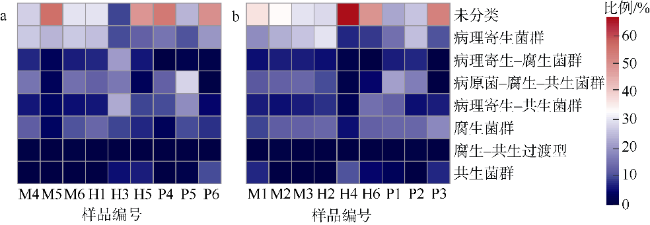
文章以丛生盔形珊瑚Galaxea fascicularis为研究目标, 对其在健康和白化两种状态下的虫黄藻密度、光合生理指标和共生真菌群落进行分析比较。结果发现白化状态下的虫黄藻密度和最大光化学量子产量(Fv/Fm)比健康状态下显著降低了75%和16%。健康和白化状态下珊瑚的共附生真菌均以子囊菌门Ascomycota、担子菌门Basidiomycota、壶菌门Chytridiomycota、毛霉门Mucoromycota为主, 但健康状态下共生真菌的Shannon多样性指数稍低于白化状态下的。健康珊瑚的虫黄藻密度受到热应激威胁而显著降低, 但仍然在合适的共生体密度范围之内。尽管白化珊瑚共生虫黄藻密度降低, 但其光合电子传递系统并没有完全遭到破坏。白化珊瑚真菌的多样性指数、真菌病原体类群及致病功能未显著增加, 显示热胁迫下珊瑚共生真菌群落的稳定性可能在热应激反应中发挥着重要作用。
朱文涛 , 夏景全 , 刘相波 , 尹洪洋 , 朱铭 , 任瑜潇 , 谢敏睿 , 黄建中 , 李秀保 . 丛生盔形珊瑚光合生理及共生真菌群落分析[J]. 热带海洋学报, 2022 , 41(2) : 132 -141 . DOI: 10.11978/2021064
Differences in zooxanthellae density, photosynthetic physiological indicators, and symbiotic fungal communities between healthy and bleaching states of Galaxea fascicularis were investigated in this study. The results showed that the zooxanthellae density and maximum photochemical quantum yield (Fv/Fm) of Galaxea fascicularis in the bleaching state were significantly lower by 75% and 16%, respectively, than those in the healthy corals. The composition of endolithic fungi community was mainly Ascomycota, Basidiomycota, Chytridiomycota, and Mucoromycota. The Shannon diversity index of endolithic fungi in healthy corals was slightly lower than that in bleaching corals, and the endolithic fungi compositions of these two coral groups had some similarity. Zooxanthellae content of healthy corals was significantly reduced due to the threat of heat stress, but it was still within the recommended appropriate symbiont density range. Despite the significant lost of density of symbiotic zooxanthellae from bleached corals, their Photosystem Ⅱ complexes were not completely destroyed. The results showed that there was no significant increase in diversity index, fungal pathogen group and predictive function in bleaching corals. The stability of endolithic fungi community may play an important role in the heat stress response. In the future, we will consider combining traditional isolation and culture techniques, high-throughput sequencing, metagenomics, and other methods to comprehensively study the diversity and ecological functions of coral endolithic fungi.
表1 调查站位的地理坐标及水体环境参数Tab. 1 Geographical locations of the sampling stations |
| 美夏海域 | 黄龙海域 | 排浦海域 | |
|---|---|---|---|
| 纬度/N | 20°00′06″ | 19°55′30″ | 19°38′30″ |
| 经度/E | 109°39′05″ | 109°31′25″ | 109°07′39″ |
| 温度/℃ | 32.53±0.93 a | 31.68±1.08 a | 31.41±0.16 a |
| pH | 8.07±0.04 a | 8.07±0.05 a | 8.10±0.02 a |
| 盐度/‰ | 32.02±0.36 a | 31.77±0.18 b | 32.30±0.13 a |
| 硅酸盐/(mg·L-1) | 0.56±0.03 a | 0.57±0.05 a | 0.43±0.06 b |
| 溶解态无机氮/(mg·L-1) | 0.27±0.04 a | 0.32±0.05 a | 0.13±0.04 b |
| 磷酸盐/(mg·L-1) | 0.03±0.00 a | 0.02±0.01 a | 0.01±0.00 b |
注: 环境参数值用均值±标准误差表示, 同行不同字母表示在p<0.05水平上差异显著 |
| [1] |
黄晖, 尤丰, 练健生, 等, 2012. 海南岛西北部海域珊瑚礁造礁石珊瑚种类组成与分布[J]. 海洋科学, 36(9): 64-74.
|
| [2] |
黄玲英, 余克服, 施祺, 等, 2011. 三亚造礁石珊瑚虫黄藻光合作用效率的日变化规律[J]. 热带海洋学报, 30(2): 46-50.
|
| [3] |
李淑, 余克服, 施祺, 等, 2007. 南海北部珊瑚共生虫黄藻密度的种间与空间差异及其对珊瑚礁白化的影响[J]. 科学通报, 52(22): 2655-2662.
|
| [4] |
李淑, 余克服, 陈天然, 等, 2011. 珊瑚共生虫黄藻密度的季节变化及其与珊瑚白化的关系--以大亚湾石珊瑚为例[J]. 热带海洋学报, 30(2): 39-45.
|
| [5] |
潘子良, 余克服, 王英辉, 等, 2019. 黄岩岛潟湖珊瑚的共生虫黄藻密度及其生态意义[J]. 广西大学学报(自然科学版), 44(6): 1752-1760.
|
| [6] |
舒为, 田晓玉, 赵洪伟, 2020. 海南海口温泉真菌、细菌多样性及其环境影响因素分析[J]. 微生物学报, 60(9): 1999-2011.
|
| [7] |
王荻潇, 钟敏, 雷晓凌, 等, 2018. 1株来自丛生盔形珊瑚的球托霉鉴定及生理学特性研究[J]. 微生物学杂志, 38(3): 57-63.
|
| [8] |
徐佳, 陈彬, 雷晓凌, 等, 2011a. 丛生盔形珊瑚共附生可培养真菌多样性分析[J]. 微生物学通报, 38(8): 1193-1198.
|
| [9] |
徐佳, 雷晓凌, 佘志刚, 等, 2011b. 丛生形盔珊瑚共附生真菌的分离及其抗菌活性的筛选[J]. 中国海洋药物, 30(5): 23-28.
|
| [10] |
张丹, 蒋群, 李英新, 等, 2015. 中国南海海绵和珊瑚共附生真菌多样性及其抑菌活性研究[J]. 中国海洋药物, 34(2): 1-9.
|
| [11] |
周洁, 施祺, 余克服, 2011. 叶绿素荧光技术在珊瑚礁研究中的应用[J]. 热带地理, 31(2): 223-229.
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
ROBERTS,
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}